Le réseau Français des instituts techniques de l’agro-alimentaire

 journées du RMT Al-CHIMIE : contaminations chimiques, des chaînes alimentaires à l'exposome
journées du RMT Al-CHIMIE : contaminations chimiques, des chaînes alimentaires à l'exposome
3 et 4 avril 2024, Nantes
Au programme de ces deux journées : état des connaissances au travers de présentations par les partenaires du RMT, visite du Laberca (laboratoire d'étude des résidus et contaminants dans l'alimentation) et de la hall de technologie d'Oniris.
 4 WEBINAIRES
4 WEBINAIRES
MAÎTRISE DES DANGERS MICROBIOLOGIQUES DES ALIMENTS : MIEUX CONNAÎTRE LES OUTILS D'AIDE À LA DÉCISION BASÉS SUR LA MODÉLISATION
Avril & mai 2024
Organisés par le RMT Actia Qualima.
Mardi 2 avril 2024
Validation des mesures de maîtrise et réglementation
Vendredi 12 avril 2024
Surveillance microbiologique de la qualité des aliments et échantillonnage
Mercredi 24 avril 2024
Validation de la durée de vie microbiologique des alimentset Appréciation quantitative de l'exposition (AQE)
Jeudi 16 mai 2024
Évaluation des risques microbiologiques (AQR) et approches multicritères
Contacts : Janushan Christy, j.christy@actalia.eu,
Valérie Stahl, v.stahl@aerial-crt.com & Catherine Denis, c.denis@actalia.eu
participation du projet européen buddie-pack![]()
au salon du vrac et du réemploi
13 et 14 mai 2024, Paris
En phase avec l'application du décret 3R (réduction-réemploi-recyclage), Buddie-Pack est un projet d’économie circulaire visant à mettre en place une approche systémique pour le déploiement à grande échelle des emballages plastiques réutilisables (RPP), basée sur une démarche multidisciplinaire combinant innovations sociales, technologiques et économiques.
L'Actia, partenaire du projet sera présente sur le stand G3/G4, avec Eternity System.
Contact : Wilfrid Paget, w.paget@actia-asso.eu
![]()
FORMATION EN LIGNE SUR LA DURÉE DE VIE DES ALIMENTS DU RÉSEAU ACTIA
3e session ouverte jusqu'au 31 mai 2024
Quels sont les outils disponibles pour déterminer la durée de vie microbiologique d’un aliment ?
À quoi servent-ils et comment les utiliser ? Quel est le contexte réglementaire ?
Quels sont les outils obligatoires et ceux qui sont facultatifs ?…
Les experts en microbiologie des aliments des Instituts techniques agro-alimentaires vous proposent cette formation en ligne illustrée d’exemples concrets.
Pour chacune des sept thématiques abordées, retrouvez une vidéo, des cas pratiques, des quizz et des ressources complémentaires.
Inscription & informations complémentaires
 Journées Aliments et Santé : rencontrez aujourd’hui l’alimentation de demain
Journées Aliments et Santé : rencontrez aujourd’hui l’alimentation de demain
19 et 20 juin 2024, la Rochelle
Le Critt agro-alimentaire de La Rochelle organise la quinzième édition des Journées Aliments et Santé, convention d’affaires réunissant plus de 1 000 professionnels de l’agro-alimentaire :
- conférences sur des thématiques clés (économie circulaire, consommateurs, sobriété, ingrédients, innovation haute technologie, aliments & santé…) ;
- rendez-vous d’affaires ;
- ateliers ;
- plus de 250 nouveaux produits à découvrir ;
- une offre variée de solutions proposées par les entreprises présentes.
L’Actia et ses centres seront mis en avant à l’occasion de cette édition, qui s’inscrira dans un objectif « zéro carbone ». www.jas-larochelle.fr
Contact : Marion Bégaud, contact.jas@crittiaa.com
 Nouveau centre de recherche et innovation de l’Ifip, au service de la filière porcine
Nouveau centre de recherche et innovation de l’Ifip, au service de la filière porcine
Les outils expérimentaux de l’Institut du porc (Ifip) sont désormais regroupés au Centre international de recherche innovation (Ciri), à Romillé près de Rennes. Au sein de ce dispositif expérimental, une nouvelle plate-forme agro-alimentaire dédiée aux viandes et charcuteries, comprend un laboratoire de microbiologie P2 (pasteurienne, biomoléculaire, phages, bases de données Salmonelles et Listeria, outils de microbiologie prévisionnelle), un laboratoire de mesures technologiques P2 (challenge-test), un laboratoire de biochimie (spectrométrie de masse, chromatographie en phase liquide à haute performance), une plate-forme technologique (technologies du cuit et du sec), une salle de découpe et un scanner tomographique.
Prolongés par des mesures en grandeur nature dans les usines et ateliers professionnels, ces outils analytiques apportent un soutien indispensable aux expertises de terrain.
Contact : gilles.nassy@ifip.asso.fr
Décembre 2023

Nouveau laboratoire P2+ pour l’Adiv
Après d’importants travaux achevés en juin dernier, le nouveau laboratoire P2+ de l'Adiv, habilité à la réalisation de protocoles in situmettant en œuvre de la contamination artificielle par des bactéries pathogènes, est désormais opérationnel. Le laboratoire et l’atelier, dont la surface a été doublée, ont été réaménagés en profondeur avec l’acquisition de nouveaux équipements, dédiés à la culture des bactéries, aux fabrications pilotes reproduisant l’ensemble des opérations unitaires employées dans le secteur des produits carnés : étuvage, fumage, cuisson, refroidissement rapide, conditionnement sous vide ou sous atmosphère modifiée, conservation. Cet investissement a été réalisé avec le soutien de France Relance.
Contact : souad.christieans@adiv.fr
Décembre 2023

La plate-forme de culture en mer connectée du Ceva
Le Ceva s’est doté d’une plate-forme expérimentale de culture en mer installée sur sa concession, dans l’estuaire du Trieux (Côtes d’Armor, Bretagne). Elle permet de réaliser un suivi de la biodiversité des algues et de la qualité des eaux, dans une zone couvrant de nombreuses activités aquacoles.
Financée dans le cadre du plan de relance, les enjeux de cette nouvelle plate-forme sont multiples :
- développer des pratiques de cultures écoresponsables en phase avec les axes stratégiques des plans de développement régionaux et nationaux ;
- mettre en place une zone pilote, témoin de la biodiversité, de la ressource biologique disponible ainsi que de la qualité du milieu ;
- mettre à disposition des données environnementales hautes fréquences ;
- élargir le champ des formations actuelles aux bonnes pratiques de culture et sensibiliser à leur impact sur le milieu.
La plate-forme est accessible aux professionnels de la filière algue ou aux industriels et scientifiques souhaitant mener des travaux de recherche sur des espèces spécifiques avec le Ceva, ainsi qu’aux porteurs de projets désireux de se former aux pratiques culturales respectueuses de l’environnement. www.ceva-algues.com
Décembre 2023

Élargissement du partenariat
du RMT Actia Prot&in
Au mois de juin, les membres du RMT ont voté à l’unanimité l’entrée d’Actalia au sein du réseau, portant ainsi à onze le nombre de partenaires.
Pierre Barrucand, directeur de l’unité « Produits laitiers » d’Actalia : « En lien avec l’important développement de produits mixtes, associant protéines laitières et végétales, notre intégration au RMT Actia Prot&in permettra de fournir un accompagnement scientifique et
technique aux acteurs et industriels de la filière laitière, en lien avec les spécificités et interactions protéines végétales / protéines laitières et la valorisation des protéines végétales, comme nouveaux ingrédients techno-fonctionnels clean label dans les matrices laitières. Nous nous réjouissons de pouvoir apporter au RMT notre expertise et notre appui technique sur les différentes actions du programme de travail ».
Alain Peyron, directeur de l’Adiv : « Cet élargissement nourrira les réflexions portant sur les problématiques et verrous rencontrés dans le cadre des approches technologiques, analytiques ou en matière de formulation et de sécurisation sanitaire, en lien avec le développement de produits associant protéines végétales et protéines animales, que nous traitons dans le RMT ».
Contacts : Benjamin Buaud, b.buaud@iterg.com & valerie.scislowski@adiv.fr,
coordinateurs du RMT Actia Prot&in
Décembre 2023
![]() Bilan du deuxième symposium franco-américain sur le recyclage & la réutilisation des emballages (PPWR2) des 9/10 novembre 2023
Bilan du deuxième symposium franco-américain sur le recyclage & la réutilisation des emballages (PPWR2) des 9/10 novembre 2023
Cette rencontre qui a eu lieu au campus Agro Paris-Saclay, à Palaiseau, avec des experts en réglementation des emballages, de recyclage, de réutilisation et d’éducation a été très instructive. Le symposium a suscité une réaction très positive de la part d’un public diversifié. Les commentaires reçus, après la clôture de l’événement, reflètent les discussions et les échanges fructueux qui ont eu lieu au cours des sessions. À cette occasion, l’Actia a présenté la plate-forme de formation européenne sur l’emballage « Fitness».
Les points forts de l’événement :
- 6 sessions uniques se sont déroulées, avec un total de 23 présentations et deux « wrap-up » de parties prenantes aux États-Unis, dans l’UE, au Chili et en Chine ;
- 167 personnes inscrites, dont 44 sur place ;
- la première journée, consacrée aux réglementations de l’UE, des États-Unis, du Chili et de la Chine ainsi qu’au recyclage, a réuni 144 participants ;
- le deuxième jour, dédié à la réutilisation et à l’éducation en matière d’emballage, a rassemblé 75 participants.
Contact : Phuong-Mai Nguyen - phuong-mai.nguyen@lne.fr
Décembre 2023
![]() webinaire du 16 octobre 2023 relatif au règlement sur les emballages et les déchets d’emballages
webinaire du 16 octobre 2023 relatif au règlement sur les emballages et les déchets d’emballages
Les cinq vidéos extraites du dernier webinaire organisé dans le cadre du projet européen Erasmus + Fitness 2 coordonné par l’Actia, sont en ligne sur la chaîne Youtube, afin de (re)découvrir les présentations réalisées sur l’économie circulaire des emballages plastiques, le contexte réglementaire du futur règlement européen sur les emballages et les déchets, ainsi que le compostage des plastiques.
Les enregistrements et supports sont également disponibles sur le site internet de Fitness 2.0.
Contact : Wilfried Paget, w.paget@actia-asso.eu
Décembre 2023
Actia guide transformation produits Bio - édition 2023
Au fil des évolutions réglementaires et depuis ses débuts, l’agriculture biologique a fait l’objet de nombreuses améliorations pour toujours mieux s’adapter aux attentes des consommateurs, en préservant ses valeurs d’origine. Dans ce contexte évolutif, les transformateurs ont besoin d’être accompagnés pour démarrer une gamme de produits Bio ou approfondir leurs connaissances sur la transformation des produits issus de l’agriculture biologique.
Cet ouvrage, fruit d’un travail collectif des membres du RMT Actia TransfoBio, est un véritable outil opérationnel destiné aux entreprises. Vous aurez accès à un autodiagnostic comportant les principales questions qu’il faut se poser au moment de se lancer dans la transformation de produits biologiques. Les éléments clés de la transformation Bio sont ensuite détaillés, en s’appuyant sur les spécificités de la règlementation Bio : marché, démarches de certification, approvisionnement, formulation, transformation. Des cas concrets issus d’entreprises, des outils pratiques et de nombreuses données sont fournis au fil de cet ouvrage. TÉLÉCHARGEZ LE GUIDE
Septembre 2023

L'ACTIA ACCOMPAGNE LES ENTREPRISES AGRO-ALIMENTAIRES POUR ASSURER L'INDUSTRIALISATION DE LEURS INNOVATIONS
Nos experts ont été sélectionnés par Bpifrance pour accompagner les entreprises agro-alimentaires dans le cadre du diagnostic « Amorçage industriel ». Celui-ci a pour objectif de soutenir dans leur projet d’industrialisation les start-up et les entreprises souhaitant se diversifier, en privilégiant les options industrielles françaises. Il permet à l’entreprise de définir sa stratégie industrielle, puis d’affiner le projet, afin de mieux préparer l’industrialisation opérationnelle et de prendre des décisions éclairées.
Prix : 10 000 € HT maximum, pris en charge à 80 % par Bpifrance.
DEMANDEZ VOTRE DIAG « AMORçAGE INDUSTRIEL »
Contact : Alice Dulas, a.dulas@actia-asso.eu
Juillet 2023
 ALIMENTATION ET SANTÉ : L'ANSES ET L'ACTIA ENGAGENT UN PARTENARIAT
ALIMENTATION ET SANTÉ : L'ANSES ET L'ACTIA ENGAGENT UN PARTENARIAT
Pr Benoît Vallet, directeur général de l’Anses, et André Pouzet, président de l’Actia, ont signé un accord de partenariat le 26 juin. Ils s’engagent ainsi à renforcer leurs coopérations pour surveiller et prévenir les risques sanitaires actuels ou émergents dans les filières alimentaires, et pour favoriser l’amélioration de la qualité des aliments.
Communiqué de presse
Juin 2023
 Webinaire « Affichage environnemental des produits alimentaires : où en est-on ? » Jeudi 20 avril 2023 - VOIR LE REPLAY
Webinaire « Affichage environnemental des produits alimentaires : où en est-on ? » Jeudi 20 avril 2023 - VOIR LE REPLAY
Les partenaires du RMT Actia Écoval travaillent sur le développement méthodologique propre à l’évaluation environnementale des produits et procédés alimentaires et sur l'information au consommateur. Ils vous proposent de faire le point sur le projet d’affichage environnemental des produits alimentaires et ce que son déploiement, prévu prochainement, implique pour les entreprises agro-alimentaires.
https://www.actia-asso.eu/projets/ecoval-2020/
Mai 2023

Audition de l’ACTIA par l’OPECST sur l’alimentation dite « ultra-transformée »
Didier Majou, directeur de l’Actia, a été auditionné le 21 octobre 2022 par l’Office parlementaire d’évaluation des choix scientifiques et technologiques (OPECST). Madame la sénatrice Angèle Préville, vice-présidente de l’OPECST, accompagnée d’un conseiller scientifique, a souhaité, en tant que rapporteur d’une note scientifique sur l’alimentation dite « ultra-transformée », recueillir le point de vue de l’Actia sur ce concept, la classification Nova et les thématiques en lien avec ces thèmes. D’autres experts et structures ont également été entendus dans le cadre de ces auditions.
Voir la note scientifique diffusée le 26 janvier dernier
Contact : Didier Majou, d.majou@actia-asso.eu
Février 2023
L’ACTIA renouvelée à la présidence de la Commission Afnor V01C « Sécurité des aliments - Hygiène et Management » Anne-Gaëlle Mellouet (CTCPA) a été reconduite dans le poste de présidente de la commission Afnor V01C, sous mandat de l’Actia, jusqu’au 14 décembre 2025. Cette commission a en charge la traçabilité et la sécurité des aliments, en termes de management et d’hygiène (terminologie, bonnes pratiques, outils analytiques), pour s’assurer de la maîtrise des conditions de fabrication à chacune des étapes de la chaîne « de la fourche à la fourchette ».
Anne-Gaëlle Mellouet (CTCPA) a été reconduite dans le poste de présidente de la commission Afnor V01C, sous mandat de l’Actia, jusqu’au 14 décembre 2025. Cette commission a en charge la traçabilité et la sécurité des aliments, en termes de management et d’hygiène (terminologie, bonnes pratiques, outils analytiques), pour s’assurer de la maîtrise des conditions de fabrication à chacune des étapes de la chaîne « de la fourche à la fourchette ».
Les Centres Actia experts en sécurité des aliments sont très impliqués dans la rédaction de ces normes, tant au niveau national qu’international.
Contact : Anne-Gaëlle Mellouet, agmellouet@ctcpa.org
Février 2023

LES EXPERTS ACTIA mobilisés pour améliorer la performance environnementale des PME agro-alimentaires
L’Ademe et Bpifrance lancent le Diag Éco-conception, en collaboration avec le pôle Éco-conception, afin de soutenir les entreprises françaises dans leur transition énergétique et écologique. L’accompagnement est mis en œuvre sur six à huit mois grâce à l’intervention d’experts qualifiés, dont les experts Actia pour le secteur agro-alimentaire.
Il permet de :
- améliorer la performance environnementale de produits, services ou procédés grâce à une évaluation environnementale ou à une analyse des pratiques ;
- former les équipes à l’éco-conception ;
- intégrer la démarche d’éco-conception dans la stratégie d’entreprise.
DEMANDEZ VOTRE DIAG « ÉCO-CONCEPTION »
Contact : Alice Dulas, a.dulas@actia-asso.eu
Janvier 2023
LES INSTITUTS TECHNIQUES AGRO-INDUSTRIELS (ITAI) ET L’ACTIA QUALIFIÉS PAR LE MINISTÈRE DE L’AGRICULTURE ET DE LA SOUVERAINETÉ ALIMENTAIRE POUR cinq ANS
Selon l’arrêté du 22 décembre 2022, le ministère de l’Agriculture et de la Souveraineté alimentaire vient d’attribuer la qualification d’Institut technique agro-industriel (ITAI) à quinze Instituts et à l’Actia, comme structure de coordination nationale des ITAI, ainsi qu’à dix-neuf Instituts techniques agricoles (ITA) fédérés par l’Acta, pour une nouvelle période de cinq ans (2023-2027). ADRIANOR est pour la première fois qualifié en tant qu’Institut technique agro-industriel, les autres bénéficient d’un renouvellement ; cinq Instituts techniques obtiennent la double qualification agricole et agro-industrielle : CEVA, IFIP, IFPC, IFV et ITAB.
Ces qualifications confirment la reconnaissance par l’État de la qualité des métiers et missions de ces organismes auprès des entreprises, pour renforcer leur compétitivité par l’innovation, la qualité et la performance.
Contact : Alice Dulas, a.dulas@actia-asso.eu
Janvier 2023

Projet Erasmus + FitNESS 2.0 sur les emballages alimentaires : nouveau site internet
Le projet Fitness 2.0, coordonné par l’ACTIA et le LNE, a pris la suite du projet Fitness (septembre 2017 - août 2020), qui avait permis de développer une plate-forme d’apprentissage en ligne sur les emballages alimentaires, à destination de l’enseignement supérieur et des entreprises. Fitness 2 touche un public plus large, de l’industrie chimique aux organismes recycleurs, en intégrant les fabricants et transformateurs d’emballage et propose des formations certifiées, validées par des tests en ligne. Le nouveau site internet regroupe toutes les informations sur le projet, les partenaires, la plate-forme d’apprentissage, ainsi que des actualités sur l’emballage alimentaire. Une section spécialement dédiée aux industriels est en cours d’élaboration.
Contact : Gemma Cornuau, g.cornuau@actia-asso.eu
Novembre 2022
 Matériaux recyclés et emballage alimentaire : le CTCPA, INRAE, IPC et le LNE renforcent leur collaboration
Matériaux recyclés et emballage alimentaire : le CTCPA, INRAE, IPC et le LNE renforcent leur collaboration
Le CTCPA, INRAE, IPC et le LNE viennent de signer un accord-cadre sur l’évaluation des risques sanitaires liés à l’utilisation de matériaux recyclés pour l’emballage alimentaire.
Ce groupe multidisciplinaire représentatif des acteurs de la filière emballage alimentaire, mobilisera les connaissances et expertises au meilleur niveau, pour appréhender les multiples conséquences de la généralisation de l’emploi de matériaux recyclés dans l’emballage alimentaire.
- Mise à disposition de méthodes d’évaluation de la sécurité des matériaux recyclés.
- Émergence de protocoles et de matériaux de référence.
- Contribution au développement de l’économie circulaire et de la réduction des déchets.
Cette collaboration répond à l’appel à manifestation d’intérêt « Emballages et contenants alimentaires et politiques de durabilité : nouvelles contraintes d’alimentarité et impacts sur la nutrition et la santé », qui vient de paraître sous l’égide de l’ANR.
Contacts : Patrice Dole, pdole@ctcpa.org - catherine.loriot@lne.fr - colette.breysse@ct-ipc.com
Novembre 2022

ITAINNOV 2022 : découvrez l’Édition spéciale !
Elle retrace les temps forts de la quatrième édition du concours d’innovations des Instituts techniques agricoles et agro-industriels, dont le point final a été la cérémonie de remise des trophées, le 2 mars 2022, au Sial. Vous y trouverez des extraits de la cérémonie, la présentation des quatre lauréats, du prix spécial du public et du jury Itainnov 2022, le témoignage d’Instituts récompensés lors des éditions précédentes et l’accès aux vidéos réalisées.
Plus d’informations - Téléchargez l’édition spéciale
Contact : Alice Dulas, a.dulas@actia-asso.eu
Novembre 2022
 Suite à l’avis du cONSEIL SCIENTIFIQUE ET TECHNIQUE de l’Actia, le ministère de l’Agriculture et de la Souveraineté alimentaire a agréé deux UMT à partir de janvier 2023, pour 5 ans.
Suite à l’avis du cONSEIL SCIENTIFIQUE ET TECHNIQUE de l’Actia, le ministère de l’Agriculture et de la Souveraineté alimentaire a agréé deux UMT à partir de janvier 2023, pour 5 ans.
UMT ACTIA QUALIVEG
« Conception intégrée des itinéraires de transformation des fruits & légumes » portée par le CTPCA et Inrae.
Contact : Stéphane Georgé, sgeorge@ctcpa.org
UMT ACTIA PROFEEL
« Lipides et protéines végétales pour des formules nutritionnelles biodisponibles et écoresponsables » portée par l’Iterg et le CRPP (Centre de recherche Paul Pascal, UMR CNRS - université de Bordeaux).
Contact : Cécile Joseph, c.joseph@iterg.com
Novembre 2022

Ce diagnostic vous permet de :
- mesurer les émissions de gaz à effet de serre de votre entreprise ;
- élaborer votre plan d’actions pour décarboner votre entreprise ;
- mettre en place les premières actions de la conduite du changement en interne et avec vos principaux clients-fournisseurs.
Cinq objectifs :
- diminution de la consommation d’énergie des entreprises ;
- réduction de la dépendance et donc de la vulnérabilité, aux énergies fossiles ;
- montée en compétences de l’entreprise sur la maîtrise de son empreinte carbone ;
- sensibilisation de vos salariés ;
- amélioration de l’image de votre entreprise.
DEMANDEZ VOTRE DIAG « DÉCARBON’ACTION »
Contact : Alice Dulas, a.dulas@actia-asso.eu
Novembre 2022

Les Centres Actia certifiés Qualiopi
Pour la plupart organismes de formation dans le domaine agro-alimentaire, les Centres Actia se sont mobilisés afin d’obtenir leur certification Qualiopi. Cette certification répond aux exigences du Référentiel national qualité. C’est une véritable reconnaissance du haut niveau de qualité des processus de formation de nos centres. Depuis janvier 2022, elle est nécessaire pour que les entreprises et les particuliers puissent faire prendre en charge les coûts des formations par des fonds publics ou mutualisés (financement par un opérateur de compétences - Opco, Pôle emploi…).
Avril 2022

L’ACTIA et INRAE signent un accord-cadre pour renforcer leur coopération
Face aux enjeux auxquels le secteur est durablement confronté, l’ACTIA et INRAE veulent renforcer leurs synergies et leur efficacité en mettant en œuvre un accord-cadre de coopération pour 5 ans.
Les deux organismes complémentaires s’engagent ainsi à traiter conjointement des questions transversales aux filières et aux disciplines à forte sensibilité sociétale. Les enjeux de sécurité sanitaire, de qualité nutritionnelle et gustative, de l’impact environnemental et de la performance des entreprises agro-alimentaires seront au cœur des échanges.
Communiqué de presse
Avril 2022

ITAINNOV 2022 : cérémonie de remise des prix LE 2 MARS au Salon International de l'Agriculture sur le stand du Ministère de l'Agriculture et de l'Alimentation
Les lauréats de chaque catégorie sont :
- « ENVIRONNEMENT - BIODIVERSITÉ - CLIMAT » : NOUVELLE MÉTHODE DE POLLINISATION par un insecte endémique de la Réunion. Un projet porté par Jean-Sébastien Cottineau de L’Armeflhor.
- « PERFORMANCE ÉCONOMIQUE & SOCIÉTALE » : EXOSCARNE® : un exosquelette qui réduit les efforts musculaires et la pénibilité du travail en abattoir. Un projet porté par Matthieu Alric de l'Adiv et développé dans le cadre de l'UMT Actia Agrobergo.
- « SÉCURITÉ SANITAIRE - SANTÉ DES PLANTES & DES ANIMAUX » : E-BROILER TRACK® : outil d’alerte du bien-être et de la santé des poulets d’élevage de chair. Un projet porté par Pauline Créach de l’Itavi.
- « LABELS & QUALITÉ DES PRODUITS » : ESTOGEL M® : gélifiant éco-responsable d’origine naturelle pour la cosmétique. Un projet porté par Guillaume Chollet d'Iterg.
- « PRIX SPÉCIAL DU PUBLIC » avec plus de 680 votes : BIOCLIMSOL® : outil d’aide à la décision pour le reboisement par des essences adaptées au changement climatique. Un projet porté par Jean Lemaire du CNPF.
Au total, plus de 11 100 vues des 12 vidéos des innovations nommées ont été enregistrées !
Mars 2022
ITAINNOV 2022 : L’ACTA & L’ACTIA DÉVOILENT LES 12 NOMMÉS DU CONCOURS ET LANCENT LE VOTE DU PUBLIC POUR ATTRIBUER LE PRIX SPÉCIAL
Parmi les 30 candidatures déposées, 12 ont été sélectionnées par le jury. Diversifiées et exemplaires, ces candidatures mettent en lumière la recherche scientifique, l’innovation et le savoir-faire des Instituts techniques agricoles et des Instituts techniques agro-industriels, dans quatre catégories distinctes. Parmi les 12 nommés, seul un projet par catégorie sera primé et recevra un trophée au salon international de l’Agriculture (SIA) le 2 mars prochain à Paris.
Pour la première fois, jusqu’au 1er mars 2022, le public aura l’opportunité de voter pour attribuer le prix spécial du public, choisi en fonction de l’impact positif de l’innovation et de sa contribution à  l’agriculture et à l’alimentation sûres, saines et durables, que l’internaute perçoit au travers de la vidéo de présentation. Pour voter, rendez-vous sur le site ITAINNOV pour découvrir les 12 nommés en image et cliquez sur « J’aime » directement sur la vidéo.
l’agriculture et à l’alimentation sûres, saines et durables, que l’internaute perçoit au travers de la vidéo de présentation. Pour voter, rendez-vous sur le site ITAINNOV pour découvrir les 12 nommés en image et cliquez sur « J’aime » directement sur la vidéo.
Janvier 2022
L’ACTIA a participé à la présentation des stratégies d’accélération

Agricoles et alimentaires du 4e plan d’investissements d’avenir
Mme Frédérique Vidal, ministre de l’Enseignement supérieur, de la Recherche et de l’Innovation, M. Julien Denormandie, ministre de l’Agriculture et de l’Alimentation, Mme Agnès Pannier-Runacher, ministre déléguée chargée de l’Industrie, en présence de M. Guillaume Boudy, secrétaire général pour l’Investissement, ont présenté les stratégies d’accélération agricoles et alimentaires du 4e plan d’investissements d’avenir (PIA4) lors d’un déplacement dans la Drôme le 5 novembre dernier.
André Pouzet, président de l’ACTIA, et Alain Peyron, président du Comité des directeurs ACTIA et directeur de l’ADIV, étaient présents. Alain Peyron a participé à la table ronde « Stratégies d’accélération au service de la 3e révolution agricole et de l’alimentation santé sur le thème des protéines.
Au total, ce sont 877,5 millions d’euros qui seront déployés sur cinq ans autour de deux stratégies d’accélération détaillées sur cette page.
Novembre 2021
ITAINNOV 2022 : l’ACTIA s’associe à l’ACTA pour lancer la quatrième édition du concours
Ce concours bisannuel valorise les initiatives et innovations techniques au profit des filières agricoles et agro-alimentaires. Pour cette nouvelle édition, le concours met en avant les impacts positifs recherchés en termes économique et sociétal, environnemental et climatique, sur la sécurité sanitaire et la santé des plantes et des animaux, ainsi que sur la qualité des produits. Les projets sont évalués et analysés par un jury composé d’experts présidé par Catherine Geslain-Lanéelle. À ses côtés, dix-huit personnalités du monde agricole et agro-alimentaire ont examiné les trente candidatures. La remise des trophées se déroulera le 2 mars 2022, au Salon international de l’Agriculture à Paris. Découvrez le livret des 30 candidatures et les interviews des porteurs de projets sur www.itainnov.com.
Octobre 2021
![]()
FORMATION EN LIGNE SUR LA DURÉE DE VIE DES ALIMENTS DU RÉSEAU ACTIA
DEUXIÈME SESSION - DU 14 OCTOBRE AU 31 DÉCEMBRE 2021
Quels sont les outils disponibles pour déterminer la durée de vie microbiologique d’un aliment ?
À quoi servent-ils et comment les utiliser ? Quel est le contexte réglementaire ?
Quels sont les outils obligatoires et ceux qui sont facultatifs ?
Les Instituts techniques agro-industriels du RMT Actia Qualima, experts en microbiologie des aliments, vous proposent cette formation en ligne illustrée d’exemples concrets pour répondre aux questions relatives à la durée de vie microbiologique, que se posent le fabricant et le conditionneur final lors de la commercialisation de produits alimentaires.
Formez-vous avec notre réseau d’experts !
Une première session réussie : plus de 80 % de satisfaction !
Un large public : entreprises agro-alimentaires et leurs fournisseurs, laboratoires, consultants qualité, enseignement technique et supérieur, pouvoirs publics… Les plus :
« La clarté des vidéos et l’accès au texte de la vidéo »
« Les ressources mises à disposition en complément de la formation »
« Les cas pratiques permettant d’apprécier des situations concrètes »
Octobre 2021

Dans la continuité du programme Agribalyse, l’ADEME, INRAE, l’ACTA, le Réseau des Instituts techniques agricoles (ITA), et l’ACTIA, le Réseau des Instituts techniques agro-alimentaires (ITAI), ont signé le 29 septembre 2021, à Paris, la création d’un Groupement d’intérêt scientifique (GIS) « Réseau pour l’évaluation environnementale des produits agricoles et alimentaires - REVALIM ». Les deux têtes de réseau et leurs onze instituts techniques agricoles et agro-alimentaires : ACTALIA, ARVALIS, CTCPA, CTIFL, IDELE, IFIP, IFV, ITAB, ITAVI, ITERG, et TERRES INOVIA, se sont engagés à renforcer leur partenariat avec l’ADEME et INRAE pour contribuer à l’élaboration des méthodes et des données permettant d’accompagner les acteurs des filières agricoles et alimentaires dans leurs démarches d’éco-conception des modes de production, de transformation et de distribution des produits. Les partenaires apporteront les outils d’aide à la décision à la hauteur des enjeux environnementaux, afin de permettre le développement de supports fiables pour l’information des consommateurs. Communiqué de presse
Octobre 2021
![]() L’ACTIA ACCOMPAGNE LES ENTREPRISES AGRO-ALIMENTAIRES POUR REFORMULER LEURS PRODUITS AU NATUREL :
L’ACTIA ACCOMPAGNE LES ENTREPRISES AGRO-ALIMENTAIRES POUR REFORMULER LEURS PRODUITS AU NATUREL :
DES LISTES D’INGRÉDIENTS SIMPLIFIÉES, DES PRODUITS MOINS TRANSFORMÉS
Ce programme s’adresse aux entreprises agro-alimentaires, qui souhaitent faire évoluer leurs gammes de produits afin de répondre aux nouvelles attentes des consommateurs.
Coordonné par Agria Grand Est et l’Actia, ce programme en trois étapes est proposé dans les régions Auvergne-Rhône-Alpes, Île-de-France, Nouvelle-Aquitaine, Grand Est, Hauts-de-France et Provence-Alpes-Côte d’azur. Plus d’information et contacts en régions.
Juillet 2021
 lancement de l’outil OPTINUT pour la formulation nutritionnelle
lancement de l’outil OPTINUT pour la formulation nutritionnelle
Il a été conçu par le Réseau Actia Nutriprevius pour permettre aux acteurs de l’agro-alimentaire d’améliorer facilement le Nutri-Score® de leurs produits en optimisant leurs recettes sous l’angle nutritionnel, grâce à la programmation linéaire. L’outil permet notamment d’intégrer des contraintes sur les quantités d’ingrédients, leur prix et les teneurs en nutriments, afin d’identifier rapidement de nouvelles recettes adaptées aux contraintes.
En savoir plus et télécharger l’outil.
Juillet 2021

Vient de paraître : Guide sur lA FOOD SAFETY CULTURE rédigé par le réseau actia référentiels
Afin d'accompagner les entreprises dans le déploiement de la « Food safety culture » ou « Culture de la sécurité des aliments », les spécialistes du Réseau Référentiels ont conçu un guide présentant l'historique du concept, ses différentes composantes et proposent aux entreprises une auto-évaluation et des pistes d’actions pour mettre en œuvre et promouvoir cette approche positive du management de la sécurité des denrées alimentaires.
Juin 2021
RITTMO AGROENVIRONNEMENT rejoint le réseau ActiA

Depuis début 2021, le centre de recherche créé en 2001 et spécialisé dans les domaines de la fertilisation, des bio-intrants et de l’agro-environnement a intégré le réseau Actia en tant que partenaire technique. Rittmo Agroenvironnement a été créé autour d’une idée novatrice, qui est de recycler intelligemment les matières organiques des sous-produits et coproduits, pour en faire des fertilisants utiles aux plantes et aux sols, dans le cadre d’une économie circulaire durable. En intégrant le réseau Actia, Rittmo souhaite développer cet aspect en se focalisant sur les sous-produits et coproduits des industries alimentaires et optimiser leur retour sur les sols agricoles dans des conditions durables.
Mai 2021

L’ACTIA accompagne les entreprises agro-alimentaires
pour réaliser des économies, gagner en rentabilité et
réduire les impacts environnementaux
L’ACTIA participe à l’opération « Diag Éco-Flux », un programme d’accompagnement d’entreprises
opéré par Bpifrance en partenariat avec l’Ademe pour mettre en œuvre des actions d’optimisation de leurs flux d’énergie, d’eau, de matières et de déchets, tout en réduisant leur impact sur l’environnement.
Huit experts Actia ont été agréés par Bpifrance et l’Ademe afin d’apporter leur expertise aux entreprises agro-alimentaire, pour identifier leurs sources d’économies et proposer un plan d’actions adapté. À l’issue du diagnostic, les entreprises sont suivies pendant une année avec des points réguliers et un bilan final. Le plan d’actions mis en place vise une rentabilité à court terme.
Contact : Alice Dulas, a.dulas@actia-asso.eu
Avril 2021

Vient de paraître : Guide sur les bonnes pratiques de prélèvement de surfaces en industrie agro-alimentaire, rédigé par lE RMT Actia Chlean
Afin d’assurer une bonne maîtrise sanitaire de l’hygiène dans les environnements industriels, il est indispensable de mettre en place des procédures de nettoyage et désinfection. La performance de ces opérations conditionne la sécurité sanitaire des denrées alimentaires produites. L’efficacité doit être mesurée L’efficacité doit être mesurée en différents points : propreté visuelle, microbiologique et chimique. Afin d’accompagner les professionnels, les partenaires du RMT Actia Chlean ont rédigé un guide regroupant des recommandations et des informations sur les différentes techniques de prélèvement de surface. Disponible en français et en anglais.
Avril 2021

![]() Actalia analyse les eaux usées pour la surveillance de la circulation de l’épidémie Covid-19 dans la population en France
Actalia analyse les eaux usées pour la surveillance de la circulation de l’épidémie Covid-19 dans la population en France
Actalia développe depuis plus de quinze ans une expertise reconnue au niveau européen en virologie des aliments dans le cadre de l’UMT Actia Virocontrol. Cette reconnaissance a permis à Actalia d’intégrer, en septembre 2020, le réseau national Obepine. Ce réseau est un consortium de recherche, créé en mars 2020, qui a pour mission d’analyser les teneurs en génomes du SARS-CoV-2 dans les eaux usées brutes afin de les utiliser comme outil de surveillance de l’épidémie de Covid-19 en France. L’un des enjeux du réseau Obepine est de suivre de manière anticipée et localement la circulation du virus dans les populations rattachées à un réseau de collecte des eaux usées, mais aussi d’anticiper sur les vagues épidémiques lorsque le virus circule à bas bruit dans les populations.
Contact : Nicolas Boudaud, n.boudaud@actalia.eu
Avril 2021
![]() Plateforme LEIA au LNE : une plate-forme inédite pour l’évaluation de l’intelligence artificielle
Plateforme LEIA au LNE : une plate-forme inédite pour l’évaluation de l’intelligence artificielle
Le plan de relance annoncé par l’État prévoit actuellement une enveloppe de 2,7 milliards d’euros de soutien au développement de marchés clés, dont celui de l’intelligence artificielle. Mais le développement de ce marché ne pourra se faire que si les systèmes d’intelligence artificielle sont rigoureusement qualifiés. C’est pourquoi le LNE travaille depuis plusieurs années à développer des méthodes afin d’évaluer la performance et la sécurité de robots agro-alimentaires et bien d’autres encore. Grâce aujourd’hui au soutien de France Relance, une nouvelle étape pourra être franchie dans l’évaluation de l’intelligence artificielle en dotant la France d’une plate-forme unique au monde, LEIA (Laboratoire d’évaluation de l’intelligence artificielle), prévue pour le premier semestre 2022. Contact : guillaume.avrin@lne.fr
Avril 2021
 Iterg acquiert Improve
Iterg acquiert Improve
renforcement de la recherche dans les huiles et protéines
L'Iterg vient d’acquérir 92,2 % des parts de la SAS Improve, plate-forme européenne dédiée à la valorisation des protéines végétales. Il constitue ainsi un groupe composé du Centre technique industriel et de la SAS Improve, devenant ainsi un acteur incontournable de la recherche appliquée de la filière des huiles et protéines végétales. Cette nouvelle entité développera une recherche appliquée sur l’ensemble de ce secteur et proposera une offre complète de prestations.
Contact : Claudie Gestin, c.gestin@iterg.com
Février 2021
 SENSALG’ : Une plate-forme dédiée à l’algue dans l’agro-alimentaire
SENSALG’ : Une plate-forme dédiée à l’algue dans l’agro-alimentaire
Avec ses atouts fonctionnels, nutritionnels, sensoriels et culinaires, l’algue a des arguments à faire valoir. Sensalg’ est la plate-forme numérique destinée aux entreprises des filières agricoles, aquacoles et agro-alimentaires, créée avec le soutien de la Draaf et du conseil régional de Bretagne. Son but est d’accompagner les récoltants, producteurs, transformateurs, utilisateurs pour leur permettre d’actualiser leurs connaissances, d’innover et de profiter de tous les atouts de cette ressource qu’est l’algue marine.
Animée par le Ceva, cette plate-forme regroupe l'expertise scientifique des quatre autres centres techniques bretons, regroupés sous la bannière Act Food Bretagne (Adria, IDmer, Vegenov, Zoopole développement) et du Centre culinaire contemporain.
Février 2021
LANCEMENT DE deux NOUVEAUX RMT 2021
Le ministère de l’Agriculture et de l’Alimentation a agréé deux RMT, à partir de janvier 2021, pour cinq ans, suite aux avis du conseil scientifique et technique de l’ACTIA.
 RMT ACTIA CHLEAN « HYGIÈNE DES ÉQUIPEMENTS »,
RMT ACTIA CHLEAN « HYGIÈNE DES ÉQUIPEMENTS »,
ANIMÉ PAR LE CTCPA
Objectif : mieux comprendre les mécanismes de détection, de prévention et de correction des contaminations chimiques et biologiques des lignes de production, afin de fournir des outils d’analyse et d’aide à la décision aux industriels, dans un contexte sociétal et environnemental fort.
Contact : Catherine Stride, cstride@ctcpa.org
 RMT ACTIA FLOREPRO « FLORES PROTECTRICES POUR LA CONSERVATION DES ALIMENTS », ANIMÉ PAR L’ADIV ET L’INRAE
RMT ACTIA FLOREPRO « FLORES PROTECTRICES POUR LA CONSERVATION DES ALIMENTS », ANIMÉ PAR L’ADIV ET L’INRAE
Objectif : travailler sur la place des flores de préservation dans la durabilité des filières (interactions dans les écosystèmes, modes d’action, traçabilité, efficacité et qualité des aliments) afin d’apporter des avancées, tant pour les professionnels que les pouvoirs publics, sur l’utilisation des agents de biopréservation dans les filières de production animales et végétales.
Contact : souad.christieans@adiv.fr
Janvier 2021
![]()
LANCEMENT DE LA FORMATION EN LIGNE DU RÉSEAU ACTIA SUR LA DURÉE DE VIE MICROBIOLOGIQUE DES ALIMENTS
Les Instituts techniques agro-industriels du RMT Actia Qualima, experts en microbiologie des aliments, vous proposent cette formation en ligne illustrée d’exemples concrets pour répondre aux questions relatives à la durée de vie microbiologique, que se posent le fabricant et le conditionneur final lors de la commercialisation de produits alimentaires.
Inscriptions dès maintenant. Formation accessible à partir du 8 décembre.
Novembre 2020
Le Critt agroalimentaire PACA vient d'obtenir 
sa certification Qualiopi
Parmi les premiers organismes de formation de l'agro-alimentaire à obtenir cette marque de qualité exigeante, cette certification sur le référentiel national unique montre d'ores et déjà le haut niveau de qualité des formations du Critt. Elle sera obligatoire au 1er janvier 2022, afin que les entreprises puissent faire prendre en charge les coûts des formations.
Cette certification qualité est délivrée au titre de la catégorie « Actions de formation »
et couvre toutes les actions collectives et individuelles.
Septembre 2020

Covid-19 - Lignes directrices pour l’adaptation des conditions de pratique de l’évaluation sensorielle
Depuis le début du déconfinement, les Centres Actia experts en évaluation sensorielle se sont associés pour rédiger des lignes directrices. Elles concernent les conditions spécifiques à mettre en œuvre pour assurer une activité d’évaluation sensorielle dans le contexte actuel de la pandémie de la Covid-19. Ces dispositions restent transitoires. Elles évoluent au fur et à mesure que les restrictions sont levées.
Ces lignes directrices concernent les tests réalisés en laboratoire. Les méthodologies mises en œuvre habituellement restent inchangées. Mais des précautions supplémentaires doivent être prises, en conformité avec les recommandations et exigences réglementaires.
Télécharger le document
Contact : Virginie Herbreteau, v.herbreteau@actalia.eu
Juillet 2020
Astredhor rejoint le réseau Actia
Depuis début 2020, l’Institut technique de l’horticulture a intégré le réseau Actia en tant que partenaire technique. Astredhor conçoit et met en œuvre des programmes de recherche et d’innovation au service des professionnels de l’horticulture, de la fleuristerie et du paysage. L’Institut s’appuie sur six unités régionales composées de dix stations d’expérimentation qui portent le projet de l’institut et le déclinent au sein de leurs territoires. Il regroupe plus de cent collaborateurs et plus de mille cent adhérents. Son intérêt principal à rejoindre le réseau est lié à ses objectifs de développement dans la chimie du végétal, les biomatériaux et les fleurs comestibles.
Juin 2020

LE GUIDE ACTIA « APTITUDE DES EMBALLAGES AU CONTACT DES ALIMENTS » MIS À JOUR EN FRANÇAIS ET DISPONIBLE EN ANGLAIS
Face à un contexte réglementaire complexe et afin d’accompagner les industriels de l’agro-alimentaire dans la mise en conformité de leurs emballages, trois centres partenaires du RMT Actia Propack food (CTCPA, IPC et LNE) ont collaboré à la rédaction de ce guide pratique à l’usage des industriels.
Mis à jour récemment sur les aspects réglementaires en français, il a également fait l'objet d'une traduction en anglais dans le cadre du projet européen Fitness.
Ce guide, en versions française et anglaise, est téléchargeable gratuitement sur le site de l'ACTIA.
Avril 2020
MOOC QUALITÉ DES ALIMENTS

À nouveau en ligne du 30 mars au 29 mai 2020
Via ce MOOC de quatre semaines, l’ACTIA vous propose d’appréhender les qualités microbiologique, nutritionnelle, sensorielle et environnementale d’un aliment et la manière dont elles interagissent.
En seulement une heure par semaine, complétez vos connaissances sur des sujets fondamentaux de l’agro-alimentaire afin de vous ouvrir à de nouvelles perspectives.
mooc.actia-asso.eu
Mars 2020

Lancement de 8 nouveaux RMT 2020
Le ministère de l’Agriculture et de l’Alimentation a agréé huit RMT, à partir de janvier 2020, pour 5 ans, suite aux avis du conseil scientifique et technique de l'ACTIA :
QUALIMA « MAÎTRISE DE LA QUALITÉ MICROBIOLOGIQUE DES ALIMENTS » - Animation : AERIAL/ACTALIA
Valérie Stahl, v.stahl@aerial-crt.com - Catherine Denis, c.denis@actalia.eu
ÉCOVAL « ÉCO-CONCEPTION ET ÉVALUATION » - ITERG
Fabrice Bosque : f.boque@iterg.com
QUADIFERM « PRODUITS FERMENTÉS ET DISTILLÉS » - IFV
Marie-Charlotte Colosio, marie-charlotte.colosio@vignevin.com
PROPACK FOOD « EMBALLAGES DURABLES ET ALIMENTS » - LNE/CTCPA
Catherine Loriot, catherine.loriot@lne.fr - Patrice Dole, pdole@ctcpa.org
TRANSFOBIO « TRANSFORMATION DES PRODUITS BIO » - CRITT PACA/ITAB
Cyril Bertrand, cyril.bertrand@critt-iaa-paca.com - Rodolphe Vidal, Rodolphe.vidal@itab.asso.fr
AL-CHIMIE « CONTAMINANTS CHIMIQUES » - ACTA/ITERG/INRAE
Émilie Donnat, emilie.donnat@acta.asso.fr - Florence Lacoste, f.lacoste@iterg.com - Jean-Michel Savoie, jean-michel.savoie@inra.fr
PROT&IN « PROTÉINES VÉGÉTALES » - ADIV/ITERG
Valérie Scislowski, valerie.scislowski@adiv.fr - Carine Bonnet, c.bonnet@iterg.com
ÉCOFLUIDES « EFFICIENCE ÉNERGIE & EAU » - CRITT PACA/CTCPA
Yvan Deloche, yvan.deloche@critt-iaa-paca.com - Marie-Pierre Labau, mplabau@ctcpa.org
Janvier 2020
![]() Lancement d’une UMT 2020
Lancement d’une UMT 2020
Suite à l’avis du conseil scientifique et technique de l'ACTIA, le ministère de l’Agriculture et de l’Alimentation a agréé une UMT à partir de janvier 2020, pour 5 ans :
PROTORISK « Caractérisation et maîtrise de la contamination des aliments par les parasites protozoaires Cryptosporidium spp., Giardia duodenalis et Toxoplasma gondii ».
Elle s’inscrit dans la continuité de l’UMT Protorisk 2015 et associe, comme précédemment, Actalia et l’EA7510-Escape qui est colabellisée par les universités de Reims Champagne-Ardenne et de Rouen Normandie et qui héberge les CNR de la Toxoplasmose et CNR-laboratoire expert Cryptosporidioses. Stéphanie Lacarbona, s.lacarbona@actalia.eu
Janvier 2020


Instruction technique de la DGAL sur la durée de vie microbiologique des aliments de décembre 2019 contribution du RMT Actia Qualima
L’instruction technique DGAL/SDSSA/2019-861 a pour objectif de préciser, de façon générale, les moyens dont les exploitants du secteur alimentaire disposent pour déterminer, valider et vérifier la durée de vie microbiologique (DVM) des denrées alimentaires qu’ils fabriquent et mettent sur le marché. Les obligations réglementaires sont rappelées. Les différents outils disponibles pour la réalisation des études de DVM sont présentés. Un logigramme est proposé pour aider, d’une part, les entreprises à hiérarchiser, au cas par cas, les moyens à déployer pour valider et/ou vérifier les DVM, et, d’autre part, les inspecteurs à vérifier la pertinence des dossiers présentés par les exploitants du secteur alimentaire.
L’instruction technique définit également les modalités de validation des dossiers de durée de vie microbiologique.
Le RMT Actia Qualima a rédigé deux documents cités en référence dans l’Instruction téléchargeables sur la page du RMT Qualima 2020 :
- Lignes directrices sur la microbiologie prévisionnelle et l’interprétation des résultats.
- Note sur l’Appréciation quantitative de l’exposition et/ou des risques.
Contacts : Valérie Stahl, v.stahl@aerial-crt.com et Catherine Denis, c.denis@actalia.eu
Janvier 2020
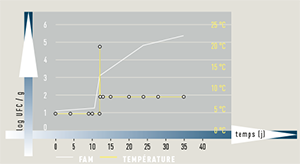
Normalisation en sécurité des aliments : le réseau Actia fortement engagé
Depuis plusieurs années, le réseau s’investit dans les activités de normalisation, tant aux niveaux national, européen, qu’international. Par son expertise reconnue en qualité et sécurité des aliments, l’ACTIA assure notamment la présidence de la commission Afnor V01C « Traçabilité et sécurité des aliments - Management et hygiène » (représentée par Anne-Gaëlle Mellouet du CTCPA), participe activement à ses groupes de travail et anime le groupe international ISO/TC 34/SC 9/WG 19 « Lignes directrices pour conduire des tests de croissance » (représentée par Florence Postollec de l’Adria). Dans ce cadre, les Centres Actia ont contribué à la parution de deux normes d’application volontaires : la norme internationale ISO 20976-1, datée de mars 2019, relative aux tests de croissance et la norme française NF V01-003, datée de décembre 2018, fixant des lignes directrices pour la réalisation de tests de vieillissement microbiologique.
À l’occasion de ces publications, la revue Lamy Droit alimentaire a interviewé Valérie Stahl et Florence Postollec, chefs de projets en microbiologie dans les ITAI Aerial et Adria, dans un article intitulé « Test de vieillissement microbiologique et test de croissance, les méthodes s’harmonisent au niveau international » dans l’édition de septembre 2019. Ces normes constituent un outil pour les exploitants du secteur alimentaire qui doivent garantir la sécurité sanitaire de leurs produits en application de la législation européenne.
Contacts : Valérie Stahl, v.stahl@aerial-crt.com et Florence Postollec, florence.postollec@adria.tm.fr
Janvier 2020
Perturbateurs endocriniens dans les contenants en PlastiqUe 
L'Assemblée nationale a créé, le 1er février 2019, une mission d'information commune sur les perturbateurs endocriniens présents dans les contenants en plastique alimentaires, cosmétiques et pharmaceutiques. Cette mission est présidée par Monsieur Michel Vialay (député des Yvelines), les co-rapporteures sont Madame Claire Pitollat (députée des Bouches-du-Rhône) et Madame Laurianne Rossi (députée des Hauts-de-Seine). L’ACTIA a été auditionnée le 11 septembre 2019 ; Didier Majou (directeur de l'Actia) était accompagné du professeur Marie-Christine Chagnon (toxicologue, Inserm, AgroSupDijon) et de Philippe Saillard (expert alimentarité des emballages, CTCPA).
L’audition a porté sur les quatre points suivants :
- déterminer les transferts de perturbateurs endocriniens présents dans les contenants en plastique vers le contenu et l’organisme humain ;
- déterminer leurs effets sur la santé et les différentes pathologies qui en découlent ;
- établir et mettre en œuvre des techniques de substitution ;
- mieux appréhender la question des transferts des perturbateurs endocriniens, issus des déchets en plastique disséminés dans la nature et les eaux usées, ainsi que leurs effets lorsqu’ils réintègrent la chaîne alimentaire.
Septembre 2019
ADRIANOR LANCE BAIA, L'ANNUAIRE DES INGRÉDIENTS ALIMENTAIRES
Forte de trente années d’expérience au service des industriels de l’agro-alimentaire, Adrianor a rassemblé au sein de l’annuaire Baia des milliers d’ingrédients, ainsi que les contacts des fournisseurs qui les proposent. Ces données sont maintenant accessibles sur le site internet d’Adrianor. Cet outil est très simple d’utilisation et facilite le quotidien des acteurs de la R&D et des achats. Il permet de mettre en relation les industriels de l’agro-alimentaire avec les fournisseurs d’ingrédients et additifs alimentaires et de rechercher des solutions alternatives à certains ingrédients pour obtenir une même fonctionnalité.
En fonction des contraintes de développement, il est possible d’affiner les résultats selon certains critères (source à privilégier, bio, vegan…).
Toutes les informations
Contact : Élodie Buchet, e.buchet@adrianor.com
Septembre 2019
LE 10 SEPTEMBRE 2019 : INAUGURATION DE LA PLATE-FORME EXPÉRIMENTALE D’IRRADIATION D’AERIAL, FEERIX®
Cette plate-forme est basée sur des technologies de pointe en matière d’accélération d’électrons et de génération de rayons X de forte énergie.
Installé dans une extension du bâtiment d’Aerial de 800 m², le Rhodotron®, qui pèse près de onze tonnes, est issu d’une technologie développée par le CEA et industrialisée par la société belge IBA, partenaire du projet; il permet d’étudier, sur une configuration originale à deux lignes de faisceaux séparées (électrons et rayons X), l’ensemble des applications multisectorielles de l’ionisation, telles que la stérilisation de dispositifs médicaux, l’amélioration des propriétés de polymères ou encore l’ionisation des aliments.
Contact : Benoît Duranton, b.duranton@aerial-crt.com
Septembre 2019
Le Cervia Paris Île-de-France devient
Île-de-France Terres de saveurs
En décidant le mercredi 19 juin 2019, lors d’une assemblée générale extraordinaire, de donner une nouvelle identité à leur organisme, les membres adhérents de l’association ont fait le choix fort de revendiquer une part importante de l’identité du territoire francilien, de la valoriser et de la faire partager par tous, professionnels des secteurs agricoles et alimentaires, ainsi qu’aux consommateurs
franciliens. Avec cette nouvelle dénomination, c’est donc l’ensemble de cette filière d’excellence francilienne allant « du champ à l’assiette », de l’amont à l’aval qui est mis en valeur de manière
distinctive et volontaire. Les missions en tant que Partenaire interface de l’ACTIA restent inchangées.
Contact : Marc Graindorge, marc.graindorge@cervia.fr
Juillet 2019
L’Itab élargit son champ d’action et devient
l’Institut de l’agriculture et de l’alimentation biologiques
 Créé pour « développer des techniques au service des systèmes agricoles Bio, productifs, durables et résilients », l’Itab élargit aujourd’hui son champ d’action « aux systèmes alimentaires Bio et durables ». Initialement Institut technique agricole, l’Itab est devenu Institut technique agro-industriel en 2017. Pour accompagner ce changement, l’Itab se nomme désormais l’Institut de l’agriculture et de l’alimentation biologiques et se dote d’un nouveau logotype. L’objectif : « accompagner plus généralement la transition vers l’agro-écologie, en cohérence avec les attentes des consommateurs et de la société », précise l’institut.
Créé pour « développer des techniques au service des systèmes agricoles Bio, productifs, durables et résilients », l’Itab élargit aujourd’hui son champ d’action « aux systèmes alimentaires Bio et durables ». Initialement Institut technique agricole, l’Itab est devenu Institut technique agro-industriel en 2017. Pour accompagner ce changement, l’Itab se nomme désormais l’Institut de l’agriculture et de l’alimentation biologiques et se dote d’un nouveau logotype. L’objectif : « accompagner plus généralement la transition vers l’agro-écologie, en cohérence avec les attentes des consommateurs et de la société », précise l’institut.
Mars 2019
MOOC QUALITÉ DES ALIMENTS
À nouveau en ligne du 7 janvier au 4 mars 2019
Via ce MOOC de quatre semaines, l’ACTIA vous propose d’appréhender les qualités microbiologique, nutritionnelle, sensorielle et environnementale d’un aliment et la manière dont elles interagissent.
En seulement une heure par semaine, complétez vos connaissances sur des sujets fondamentaux de l’agro-alimentaire afin de vous ouvrir à de nouvelles perspectives.
mooc.actia-asso.eu
Décembre 2018
Six ITAI reconnus par la DGAL pour valider les dossiers de durée de vie microbiologique des aliments
 Les exploitants du secteur alimentaire doivent s’assurer que les denrées alimentaires mises sur le marché ne sont pas dangereuses. Pour cela, ils élaborent un plan de maîtrise sanitaire afin de définir des mesures de maîtrise des dangers identifiés et vérifient que ces mesures de maîtrise sont efficaces. La validation et la vérification des durées de vie microbiologiques des aliments font partie de ces mesures de maîtrise. Certains exploitants du secteur alimentaire ou des interprofessions peuvent souhaiter bénéficier d’un avis complémentaire sur leurs dossiers de durée de vie microbiologique.
Les exploitants du secteur alimentaire doivent s’assurer que les denrées alimentaires mises sur le marché ne sont pas dangereuses. Pour cela, ils élaborent un plan de maîtrise sanitaire afin de définir des mesures de maîtrise des dangers identifiés et vérifient que ces mesures de maîtrise sont efficaces. La validation et la vérification des durées de vie microbiologiques des aliments font partie de ces mesures de maîtrise. Certains exploitants du secteur alimentaire ou des interprofessions peuvent souhaiter bénéficier d’un avis complémentaire sur leurs dossiers de durée de vie microbiologique.
Dorénavant, les expertises des ITAI membres du réseau RMT Actia Qualima, ACTALIA, ADIV, ADRIA, AÉRIAL, CTCPA, IFIP, sont reconnues par la direction générale de l’Alimentation du ministère de l’Agriculture et de l’Alimentation : les dossiers de validation de la durée de vie qu’ils rédigent sont considérés comme validés de fait par l’administration.
Avril 2018

Projet PACK_AGB : quelles performances environnementales
pour les emballages alimentaires ?
Pack_AGB a pour objectif d’améliorer l’évaluation des emballages des produits alimentaires dans la base Agribalyse. Cette base de données, propriété de l’Ademe, caractérise par l’analyse de cycle de vie (ACV) les impacts environnementaux de près de 2 500 produits alimentaires et constituera la base de référence pour le futur dispositif d’affichage environnemental en cours de développement par le ministère de la Transition écologique et l’Ademe. Ce projet s’inscrit dans la feuille de route du groupement d’intérêt scientifique Revalim qui, depuis 2021, pilote les évolutions de la base Agribalyse dans le cadre d’une gouvernance partagée entre l’Ademe, Inrae, l’Actia et l’Acta.
Il est cofinancé par l’Ademe et coordonné par le CTCPA avec Actalia, Ifip Iterg, IPC et Bleu Safran.
Prenons l’exemple d’un yaourt : chaque élément de l’emballage (pot, opercule, étiquette, fourreau), tel qu’il est vendu aux consommateurs, a été caractérisé pour une évaluation précise des impacts associés : matériaux, procédés et leurs pertes, transports et fin de vie avec les impacts évités par le recyclage. Les données produites alimenteront la prochaine version d’Agribalyse prévue mi-2024.
Contact : Margaux Colombin, mcolombin@ctcpa.org
Décembre 2023
![]() Le concept d’empreinte chimique
Le concept d’empreinte chimique
pour mieux orienter les flux de matières recyclées
Julien Kermovant a soutenu sa thèse d’université à Paris-Saclay le 21 mars 2023. Celle-ci, menée au sein de l’UMT Actia Safemat, présente une nouvelle approche basée sur une analyse originale des chromatogrammes, dans le cadre de la théorie de l’information.
Cette méthode du traitement de signal permet, sans calibration préalable, de sortir les ions les plus significatifs pour un échantillon inconnu et sans référence identifiée. L’originalité de l’approche réside essentiellement dans un alignement des codes empreintes, préalablement à leur comparaison. Cette potentialité répond à un besoin de l’industrie dans le cadre d’une économie circulaire : à partir de quand un échantillon dévie-t-il des standards de production ou de recyclage ? La réponse comprend deux niveaux :
« Quelle est l’amplitude de la différence ? », « Quelles parties du chromatogramme expliquent-elles cette différence ? »
Elle permet d’apparier les profils de contamination des matériaux et des aliments entre eux et d’orienter l’utilisation de la matière recyclée, en fonction de son niveau de décontamination reconstruit par la méthode.
Contacts : phuong-mai.nguyen@lne.fr & olivier.vitrac@agroparistech.fr
Décembre 2023
DHA (oméga-3) : un rôle clé dans l’absorption cérébrale du glucose
Les neurones présentent une forte demande énergétique et nécessitent un apport continu de glucose par le sang. En réponse à des stimuli, le métabolisme du glucose et le flux sanguin cérébral sont étroitement coordonnés pour maintenir la fonction cérébrale appropriée. L’absorption du glucose à travers la barrière hémato-encéphalique est facilitée par le transporteur GLUT-1. La première réponse aux demandes urgentes de glucose est d’augmenter le flux sanguin par vasodilatation générée par le CO₂. La seconde consiste à augmenter la densité de GLUT-1 par sa translocation à partir des réserves intracellulaires. La troisième consiste à augmenter la synthèse du GLUT-1 en stimulant la transcription du gène SLC2A1. Un tandem de deux molécules clés, l’estradiol libre et le DHA, est impliqué dans cette régulation. En particulier, il est montré qu’à partir d’un certain niveau de carence chronique en DHA, un déséquilibre permanent s’installe entre l’apport en glucose et le métabolisme cérébral. Cette carence en DHA est un facteur aggravant de certaines neuropathologies.
Didier Majou (ACTIA) & Anne-Lise Dermenghem (ACTIA)
Contact : a.dermenghem@actia-asso.eu
Décembre 2023
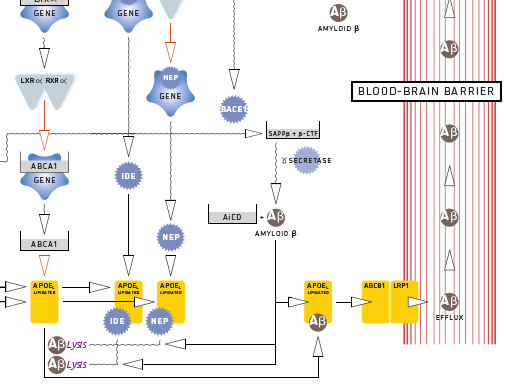
Effets du DHA (acide gras oméga-3) et de l'estradiol sur la régulation du β-peptide amyloïde dans le cerveau
Article publié dans « Brain research »
Dans les premiers stades de la maladie d'Alzheimer, il existe une forte corrélation entre les troubles de la mémoire et les niveaux corticaux d'oligomères peptidiques amyloïdes-β solubles (Aβ). Les Aβ perturbent la fonction synaptique glutamatergique, ce qui entraîne les déficits cognitifs caractéristiques de la maladie d'Alzheimer. Cet article décrit les mécanismes pathogènes qui sous-tendent l'amylose cérébrale. Ces mécanismes dépendent de la protéine précurseur de l'amyloïde et concernent la synthèse des peptides Aβ avec une compétition entre la voie non amyloïdogène et la voie amyloïdogène (entre les enzymes ADAM10 et BACE1) d'une part, et les différents processus de clairance des résidus Aβ d'autre part. Cette clairance mobilise à la fois des endopeptidases (NEP et IDE) et des transporteurs d'élimination à travers la barrière hémato-encéphalique (LRP1, ABCB1 et RAGE). L'ApoE lipidée joue également un rôle majeur dans tous les processus. La perturbation de ces voies induit une accumulation d'Aβ. La description des mécanismes révèle deux molécules clés en particulier :
- l'estradiol libre, qui a une action génomique et non génomique ;
- et le DHA en tant que ligand préférentiel des hétérodimères PPARα-RXRα et PPARɣ-RXRα.
Lorsqu'un certain niveau de carence chronique en DHA et en estradiol libre est atteint, un déséquilibre permanent s'établit dans le système nerveux central.
Didier Majou (ACTIA) & Anne-Lise Dermenghem (ACTIA)
https://doi.org/10.1016/j.brainres.2023.148681
Contact : a.dermenghem@actia-asso.eu
Décembre 2023

Le logiciel de microbiologie prévisionnelle Sym’previus intègre deux nouveaux modules sur les flores fongiques et l’emballage
Un nouvel outil pour anticiper le développement des moisissures
Le développement des moisissures est complexe car il se produit en plusieurs étapes (germination, élongation des hyphes, prolifération mycélienne, sporulation), dont les cinétiques dépendent de facteurs environnementaux comme la température, l’activité de l’eau, la présence d’acides organiques ou de composés fongistatiques ou encore la composition de l’espace de tête dans l’emballage.
Alors que la microbiologie prévisionnelle appliquée aux moisissures existe depuis de nombreuses années, aucune application visant son utilisation dans le contexte industriel n’était encore disponible. L’application en ligne Sym’Previus comporte un nouveau module fongique qui vient combler ce manque.
Un nouvel outil pour optimiser le couple emballage / atmosphère en fonction de l’aliment
En réponse à une forte demande de la part des consommateurs, les emballages sous atmosphère modifiée (MAP) permettent de prolonger la durée de vie des aliments emballés, en évitant l’ajout de conservateurs ou de stabilisants chimiques. Le nouveau module MAP facilite la sélection des emballages adéquats et des mélanges gazeux, en lien avec la formulation de l’aliment et ses conditions de stockage, pour optimiser la durée de vie des produits.
Sym’previus est le fruit d’une collaboration entre l’Actia, les ITAI (Actalia, Adiv, Adria, Aerial, CTCPA, Ifip), des entreprises agro-alimentaires (Danone, Groupe Bel, Pernod Ricard, Savencia Fromage & Dairy), une organisation interprofessionnelle (Cniel) et des laboratoires publics français (Enva, Inrae, Lubem).
Contact : Yvan Le Marc - symprevius@adria.fr - https://symprevius.eu/fr
Décembre 2023
![]() Lancement du projet Salmobond (2024-2027)
Lancement du projet Salmobond (2024-2027)
« Facteurs d’adaptation de Salmonella dans les environnements de production porcins et laitiers : développement d’outils de caractérisation moléculaire et phénotypique »
Initié au sein de l’UMT Actia Fastypers et regroupant comme partenaires deux ITAI (Actalia, Ifip-institut du porc), deux laboratoires de l’Anses (unité AB2R, unité sel), un d’Inrae (équipe B3D, institut Micalis). Ce projet financé par FranceAgriMer vise à développer :
- un outil PCR innovant, rapide et spécifique pour trois séro-vars majeurs de Salmonella afin d’identifier les marqueurs génomiques impliqués dans la formation de biofilms et dans l’adaptation et la résistance aux biocides des souches issues des filières laitière et porcine ;
- des outils phénotypiques pour caractériser les souches capables de former des biofilms et de croître à nouveau, malgré l’utilisation de biocides sur les surfaces dans les ateliers agro-alimentaires.
Ces outils seront transférés aux Instituts techniques, au service des industries agro-alimentaires, pour une utilisation simple, rapide et automatisable. Une caractérisation plus complète des souches détectées permettra ainsi aux professionnels de choisir les mesures sanitaires les plus adaptées.
Contact : Valérie Michel, v.michel@actalia.eu
Décembre 2023
 Projet Flegme : fermentation des légumes
Projet Flegme : fermentation des légumes
Ce projet participatif impliquant Vegenov et le CTCPA a exploré les communautés microbiennes, la qualité nutritionnelle et la perception par les consommateurs des légumes fermentés. La revue de littérature scientifique réalisée dans le cadre de ce projet vient tout juste d’être dévoilée dans la revue « Trends in Food Science & Technology ».
Le projet Flegme (2019-2022), piloté par Vegepolys valley avec la coordination scientifique d’Inrae et le soutien financier des régions Bretagne et Pays de la Loire, a permis de bâtir un socle solide de connaissances autour des légumes fermentés. Il a abouti notamment à de nombreux documents pratiques à l’usage des fabricants, accessibles sur le site de Vegepolys Valley, comme un ABC des légumes fermentés. Ce guide apporte notamment des éléments sur les étapes critiques d’une fermentation, les critères microbiologiques à suivre et l’étiquetage en tant que DLC ou DDM, en fonction du type de produit fabriqué.
Descriptif du projet - Les livrables
Contacts : Céline Baty-Julien, baty@vegenov.com & Stella Planchon, splanchon@ctcpa.org
Décembre 2023
Deuxième phase du projet européen « Plan P »
 Ce projet, dont l'Adria est partenaire, vise à développer un système intelligent pour la conception de nouveaux aliments durables à base de protéines végétales, type émulsion ou mousse. Basé sur l’analyse spectrale couplée à des algorithmes d’intelligence artificielle, cette solution permettra d’accélérer la transition alimentaire végétale.
Ce projet, dont l'Adria est partenaire, vise à développer un système intelligent pour la conception de nouveaux aliments durables à base de protéines végétales, type émulsion ou mousse. Basé sur l’analyse spectrale couplée à des algorithmes d’intelligence artificielle, cette solution permettra d’accélérer la transition alimentaire végétale.
Plan P a conduit au développement et à l’analyse d’une base de données de caractérisation techno-fonctionnelle d’ingrédients protéiques d’origine végétale (différentes sources / différents fournisseurs). Les ingrédients sélectionnés sont utilisés pour produire des matrices expérimentales de microstructures pour quatre domaines, produits en combinant des variables de procédés de fabrication et les contraintes de mélange. Ces matrices ont été caractérisées d’un point de vue textural. Leur signal spectral a été acquis en utilisant la spectroscopie proche infrarouge (NIR).
Les données seront traitées pour construire un modèle prévisionnel de l’acceptabilité de la texture pour les quatre domaines produits, sur la base des données spectrales acquises pendant la fabrication. En parallèle, un prototype sera testé dans un environnement de fabrication de produits agro-alimentaires pour confirmer les capacités de la solution développée.
Pour cette deuxième phase, un questionnaire est en ligne pour connaître les attentes et besoins des industriels. Ainsi l’outil développé servira concrètement dans le développement produit.
Le financement du projet est assuré par des fonds nationaux
(conseil régional de Bretagne, France - GUDP au Danemark - GSRI en Grèce) et via le programme de recherche & innovation Horizon 2020 de l’Union européenne.
Contact : jonathan.thevenot@adria.fr
Décembre 2023
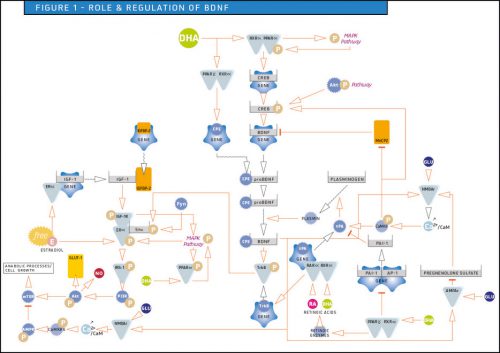
L’acide gras oméga-3 DHA augmente l'action stimulante du facteur neurotrophique dérivé du cerveau (BDNF)
Les neurones ont un besoin énergétique élevé, nécessitant un apport continu de glucose par le sang. Une régulation étroite du métabolisme du glucose, en réponse à des stimuli, est essentielle pour le bon fonctionnement du cerveau. Dans un article précédent, nous avons déjà détaillé les mécanismes par lesquels la voie de signalisation PI3K/Akt est impliquée dans l'efficacité de l'absorption cérébrale du glucose en stimulant l'action du transporteur du glucose, GLUT-1, et la vasodilatation via le monoxyde d’azote. Dans cet article, nous expliquons comment l'activation de la protéine BDNF (facteur neurotrophique dérivé du cerveau) contribue à stimuler la voie de signalisation IRS-1/PI3K/Akt et à moduler l'expression du récepteur NMDA, activé par le glutamate, qui est essentiel à la mémoire et à la plasticité synaptique. Globalement, une activité neuronale à haute fréquence induit la synthèse de BDNF et l’intervention de cette protéine dope cette voie majeure. Le DHA, comme ligand préférentiel des facteurs de transcription PPARα-RXRα et PPARɣ-RXRα, est impliqué dans la régulation critique de la stimulation par le BDNF. Le BDNF stimule la voie de signalisation IGF-1/estradiol/PI3K/Akt, et l'acide docosahexaénoïque, ou DHA, acide gras polyinsaturé oméga-3, renforce les actions du BDNF.
Contact : a.dermenghem@actia-asso.eu
D. Majou & A.L. Dermenghem,
Décembre 2023

VINGT-SIX FICHES PÉDAGOGIQUES POUR APPROFONDIR VOS CONNAISSANCES SUR L’EMBALLAGE
Les partenaires du RMT Actia Propack Food continuent à s’investir sur le développement de ressources permettant la transmission la plus large possible de leurs connaissances concernant les emballages au contact des aliments, sujet de haute importance, en particulier sur les aspects sanitaires et environnementaux.
Vingt-six fiches sont désormais disponibles en téléchargement gratuit sur leur site internet. Destinées principalement aux étudiants et enseignants (lycées techniques, BTS, DUT) ainsi qu’aux industriels, elles présentent des notions clés en science des emballages.
Juillet 2023
![]() Nouvelle parution : le changement d'échelle de la Bio
Nouvelle parution : le changement d'échelle de la Bio
Le marché des produits Bio a connu une forte croissance ces dernières années (augmentation de la demande, de l’offre, des voies de distribution…). Les opérateurs ont bénéficié de cette croissance sans toujours en connaître les causes profondes et donc sans forcément en anticiper les conséquences. Depuis 2019, les marchés Bio sont chahutés et en repli depuis 2021. Cette situation conjoncturelle peut devenir structurelle si les opérateurs ne prennent pas en charge, de façon concertée, le pilotage de leurs marchés.
Le RMT Actia Transfobio propose une analyse du changement d'échelle de la Bio et des opportunités et menaces qui se présentent aux transformateurs, afin de mettre en place les actions nécessaires pour pérenniser leurs marchés. Téléchargement gratuit
Juillet 2023
 OPTINUT - NOUVELLE VERSION 2023 DE L’OUTIL D’AIDE À LA FORMULATION NUTRITIONNELLE
OPTINUT - NOUVELLE VERSION 2023 DE L’OUTIL D’AIDE À LA FORMULATION NUTRITIONNELLE
Le Réseau Actia Nutriprevius regroupe des acteurs des Centres Actia, de la recherche et de l’enseignement autour de l’enjeu de l’amélioration de la qualité nutritionnelle pour une offre alimentaire durable. Après une première version de l’outil Optinut lancée en 2021, la version 2023 intègre le nouvel algorithme du Nutri-Score pour les aliments solides (hors boissons, matières grasses ajoutées et fromages), suite à la validation par le comité de pilotage du Nutri-Score à l’été 2022. Ces modifications doivent encore être intégrées à la réglementation nationale pour être appliquées et de cette façon, renforcer l’information des consommateurs.
Optinut permet d’intégrer des conditions sur les quantités d’ingrédients, leur prix, les teneurs en nutriments et de faciliter le décryptage du Nutri-Score et son amélioration critère par critère et, ainsi, de proposer de nouvelles recettes adaptées à l’ensemble de ces conditions.
Juillet 2023
 LANCEMENT DU PROJET ANR SUBLIM : IDENTIFICATION D’INDICATEURS POUR UNE MEILLEURE MAÎTRISE DU RISQUE DE CONTAMINATION DES ATELIERS AGRO-ALIMENTAIRES PAR LISTERIA MONOCYTOGENES
LANCEMENT DU PROJET ANR SUBLIM : IDENTIFICATION D’INDICATEURS POUR UNE MEILLEURE MAÎTRISE DU RISQUE DE CONTAMINATION DES ATELIERS AGRO-ALIMENTAIRES PAR LISTERIA MONOCYTOGENES
Initié dans le cadre du RMT Actia Chlean, ce projet est financé par l’ANR pour une durée de quatre ans et regroupe six partenaires (Anses de Boulogne-sur-Mer - coordination, Anses de Fougères, Inrae de Theix, IUT de Saint-Brieuc, Actalia et Ifip). L’objectif est d’identifier des indicateurs biologiques, moléculaires, physico-chimiques et /ou environnementaux permettant de qualifier les surfaces agro-alimentaires vis-à-vis du risque d’implantation de Listeria monocytogenes dans les filières charcuterie, produits de la mer et végétaux.
Contact : graziella.midelet@anses.fr
Juillet 2023
 LANCEMENT DU PROJET ANR SUPERSHIELD : ÉVALUATION ET OPTIMISATION DU SUPERCHILLING OU ULTRA-RÉFRIGÉRATION, PAR DES TECHNOLOGIES INNOVANTES
LANCEMENT DU PROJET ANR SUPERSHIELD : ÉVALUATION ET OPTIMISATION DU SUPERCHILLING OU ULTRA-RÉFRIGÉRATION, PAR DES TECHNOLOGIES INNOVANTES
La technologie du superchilling a un potentiel considérable : fournir un produit frais de haute qualité et d’une durée de conservation plus longue que celle des aliments réfrigérés. La température du produit doit être maintenue à environ 1 °C en dessous de la température de congélation commençante et la teneur en glace autour de 30 %. Ce projet, financé par l’ANR, a été initié au sein du RMT Actia Écofluides et regroupe Inrae (Frise, Micalis), Oniris (Gepea), Ifip, le Critt Agroalimentaire Sud, le Pôle Cristal et IMT-Atlantique. Les résultats aideront les acteurs de la filière agro-alimentaire et les responsables politiques dans leurs prises de décision concernant cette technologie, qui commence à avoir une existence légale en Europe.
Contact : yvan.deloche@critt-iaa-paca.com
Juillet 2023

L’ACTIA PARTENAIRE D’UN NOUVEAU PROJET HORIZON EUROPE : CONSERWA « VERS UNE GESTION AGRO-ÉCOLOGIQUE DES ADVENTICES DANS DIFFÉRENTS SYSTÈMES AGRICOLES ET RÉGIONS EUROPÉENNES »
Ce projet répondra à trois objectifs pour une gestion plus durable des adventices :
- mettre en place les moyens techniques permettant le développement à grande échelle des pratiques agro-écologiques et la gestion des adventices ;
- faciliter l’adoption de ces pratiques par les agriculteurs en travaillant aux niveaux des politiques, des infrastructures et de la formation ;
- montrer l’intérêt de ces alternatives non chimiques sur le changement climatique.
Coordonné par le Cyric, ce projet rassemblant vingt-trois partenaires provenant d’onze pays européens durera quatre ans, avec un budget d’environ 5 millions d’euros. L’ACTIA, ainsi que le CTCPA, apporteront leur expertise concernant l’impact de la gestion des adventices sur la qualité des produits transformés.
Contacts : Christophe Cotillon, c.cotillon@actia-asso.eu &
Gemma Cornuau, g.cornuau@actia-asso.eu
Juillet 2023
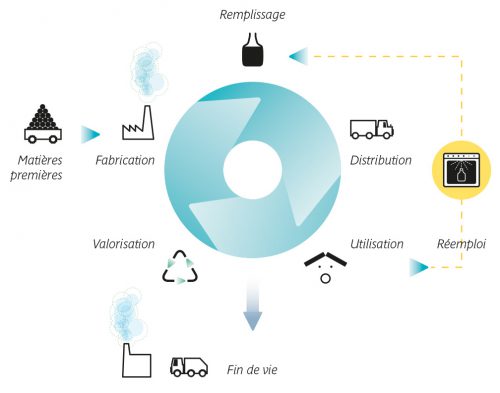 Résultats du PROJET ACV RÉEMPLOI : POUR DÉVELOPPER UNE MÉTHODOLOGIE D’ACV PROPRE AUX EMBALLAGES RÉEMPLOYABLES EN PLASTIQUE
Résultats du PROJET ACV RÉEMPLOI : POUR DÉVELOPPER UNE MÉTHODOLOGIE D’ACV PROPRE AUX EMBALLAGES RÉEMPLOYABLES EN PLASTIQUE
Avec le soutien financier de l’ACTIA, trois partenaires membres du RMT Actia Propack Food, le LNE, le CTCPA et IPC (coordinateur) ont mené le projet ACV Réemploi sur 2022. Il visait à développer une méthodologie d’ACV propre aux emballages réemployables en plastique, afin d’accompagner les entreprises dans l’évolution des législations favorables au réemploi, et à éclairer leurs prises de décision.
Résultats en fin de lettre Actialités dans le Focus R&D.
Juillet 2023
![]() L’application de microbiologie prévisionnelle Sym’previus évolue
L’application de microbiologie prévisionnelle Sym’previus évolue
Sym’previus est un outil complet de prévision des données microbiologiques, fruit d’une collaboration entre des entreprises agro-alimentaires, des organisations interprofessionnelles, des ITAI et des laboratoires publics français. Reconnu par la communauté scientifique, il permet aux industriels de garantir la sécurité et la qualité des aliments. En 2023, il sera enrichi de deux modules :
module « Fongique » : certains de ces micro-organismes fongiques peuvent présenter un risque sanitaire, et d’autres altérer les produits. Aucun outil de simulation du comportement fongique n’est aujourd’hui disponible pour les industriels. Le module « Fongique » comblera cette lacune ;
module « Map » : les emballages sous atmosphère modifiée (Map) permettent de prolonger la durée de vie en rayon des aliments emballés, sans ajout de conservateurs ou de stabilisants chimiques. Le module Map du logiciel Sym’previus a été développé pour visualiser l’impact du choix des mélanges gazeux adaptés au produit et de l’emballage, sur le développement microbiologique.
Contact : serviceclients@adria.fr
Mars 2023

NITRATES-NITRITES ET CHARCUTERIES - L’ANSES A RENDU SON VERDICT
Pour des raisons de santé publique (relation entre nitrite et cancer colorectal) et d’évolution sociétale, les tendances actuelles convergent vers une réduction des taux de nitrite et de nitrate ajoutés dans les charcuteries salaisons. La Commission européenne a entamé un projet de révision du règlement (CE) n° 1333/2008 sur les doses d’utilisation des nitrites et des nitrates. Parallèlement, le ministère de la Santé a saisi l’Anses pour évaluer le risque nitrate/nitrite dans les aliments : lire le rapport.
L’Adiv et l’Ifip avec l’appui de la Fict et de l’Actia, sont fortement impliqués pour accompagner les entreprises dans la réduction des teneurs en additifs, tout en garantissant la sécurité sanitaire. Ils sont notamment parties prenantes du « Plan d’action pour la réduction de l’utilisation des additifs nitrites/nitrates » porté par le cabinet du ministère de l’Agriculture et de la Souveraineté alimentaire. Contacts : souad.christieans@adiv.fr & gilles.nassy@ifip.asso.fr
Févier 2023
Le RMT Actia Écofluides présente une vidéo sur le Séchage par vapeur d’eau surchauffée (VES)
Des partenaires du RMT ont réalisé une vidéo expliquant le principe et les avantages du séchage par VES en industrie agro-alimentaire. Ce procédé de déshydratation repose sur l’utilisation de la vapeur d’eau surchauffée comme fluide sécheur. Il offre à l’industriel de multiples avantages : suppression de la pollution atmosphérique, amélioration de la qualité des produits et, selon le système mis en place, forte réduction des consommations énergétiques.
Contacts : yvan.deloche@critt-iaa-paca.com & Marie-Pierre Labau, mplabau@ctcpa.org
Février 2023

SOUTIEN FINANCIER DE L'ACTIA AU projet MicroPlast sur l’évaluation de méthodes de caractérisation de microplastiques en milieu marin
Dans le cadre du RMT Actia Propack Food, le LNE et ses partenaires (AgroParisTech/Inrae, Cnam/ Pimm et IPC) se sont associés à l’expédition Sea Plastics 2021 dans le cadre du projet Microplast. Pilotée par plusieurs étudiants d’AgroParisTech, cette expédition vise à sensibiliser le grand public à la problématique de la pollution marine et à contribuer à la recherche scientifique sur les micros et nanoplastiques. Elle a parcouru la mer Méditerranée de mars à juillet 2021. Avec un stock de plastique actuellement estimé à 1,2 million de tonnes, la mer Méditerranée est une zone d’accumulation importante de plastiques du fait de son caractère fermé et de son important bassin versant. L'objectif du projet Microplast a été d’inventorier et d’évaluer les méthodes existantes d’échantillonnage, de stabilisation, de préparation et de caractérisation de particules polymères à des tailles inférieures à 5 mm disséminées dans le milieu marin, en accompagnement de la campagne d’échantillonnage réalisée par Sea Plastics. Voir l'article complet dans le focus R&D de la lettre Actialités d'octobre 2022.
Novembre 2022
![]() Lancement du projet Horizon Europe BUDDIE-PACK sur les emballages plastiques réutilisables
Lancement du projet Horizon Europe BUDDIE-PACK sur les emballages plastiques réutilisables
Le projet européen Buddie-pack, financé par le programme Horizon Europe (sept millions d’euros), a démarré en septembre, pour trois ans. Coordonné par le Centre technique industriel de la plasturgie et des composites (IPC), il rassemble dix-neuf partenaires de six pays différents. L’ACTIA ainsi que le LNE et le CTCPA, sont partenaires de Buddie-pack. Ce projet d’économie circulaire a pour objectif le déploiement à grande échelle des emballages plastiques réutilisables (RPP). Il s'agira de :
- promouvoir l’acceptation des RPP auprès des consommateurs ;
- étudier les solutions de nettoyage innovantes ;
- étudier la durabilité des propriétés fonctionnelles jusqu’à trois cents cycles.
Buddie-pack devrait permettre d’éviter chaque année la production de près de 544 millions d’emballages à usage unique, de réduire la consommation d’eau et celle de plastiques vierges de 103000 tonnes et d’éviter le rejet de microplastiques dans les océans.
Contacts : Christophe Cotillon, c.cotillon@actia-asso.eu
& Gemma Cornuau, g.cornuau@actia-asso.eu
Novembre 2022
 Le CTCPA explique le concept des « hurdle technologies » dans une vidéo ludique
Le CTCPA explique le concept des « hurdle technologies » dans une vidéo ludique
Les « hurdle technologies » ou « technologies barrière », combinaison de technologies douces de préservation successives ou simultanées, constituent des alternatives crédibles aux procédés purement thermiques actuels. Elles permettent de créer de nouveaux produits ou d’obtenir une durée de consommation plus longue, d’ouvrir des pistes innovantes de consommation pour des produits peu adaptés aux procédés de conservation conventionnels et rendent possible l’utilisation d’emballages plus simplement conçus et à moindre impact environnemental.
Contact : Patrice Dole, pdole@ctcpa.org - Découvrez la vidéo
Novembre 2022

Surveillance de la dynamique des variants du SARS-CoV-2 dans les eaux usées brutes en France : nouvelle publication scientifique dans la revue « Science of The Total Environnement ».
Tout au long de la pandémie de Covid-19, de nouveaux variants ont continuellement émergé, se propageant dans les populations. Parmi ceux-ci, les variants préoccupants ont été les principaux responsables des vagues épidémiques successives, en raison de leur transmissibilité, de leur pathogénicité ou de leur capacité à échapper à la réponse immunitaire.
Cette nouvelle publication scientifique avait pour but de dater l’émergence des variants préoccupants du Sars-Cov-2, d’Alpha au sous-variant BA.2 d’Omicron, en surveillant les eaux usées de la région parisienne et de quelques villes en province. Il s’agissait aussi de modéliser la dynamique de propagation de ces variants et de caractériser la cinétique de remplacement des populations prévalentes. Ce travail a été effectué dans le cadre du réseau Obépine et de l’UMT Actia Virocontrol (Actalia, LCPME [CNRS, université de Lorraine]).
Contact : Nicolas Boudaud, n.boudaud@actalia.eu
Lire l’article
Novembre 2022
![]() Le robot Vinimag©, au service de la détection précoce des critères œnologiques des variétés d’intérêt, à l’échelle du pied de vigne
Le robot Vinimag©, au service de la détection précoce des critères œnologiques des variétés d’intérêt, à l’échelle du pied de vigne
Vinimag© est un robot de vinification de vins rouges en petit volume (<1 kg), issu de cinq ans de mise au point par l’IFV, l’Inrae et l’Institut Agro Montpellier pour respecter les grands principes de la vinification en rouge à l’échelle du kilo. Ces travaux ont été réalisés dans le cadre de l’UMT Actia Minicave (2017-2021). Ce dispositif sera utilisé dans le cadre de programmes de création et de sélection variétale dans la phase de détermination des critères œnologiques des variétés d‘intérêt, en particulier l’adaptation au changement climatique. La détection, à l’échelle du pied de vigne, des critères œnologiques des variétés d’intérêt sera réalisée et accélérée par criblage à haut débit. C’est tout l’objectif de la nouvelle UMT Actia Œnotypage lancée en janvier 2022, pilotée par l’IFV, avec l’Inrae et l’Institut Agro Montpellier. Contact : marie-agnes.ducasse@vignevin.com
En savoir plus
Novembre 2022

Clôture du projet ANR Minimeau porté par les RMT Actia Écofluides et Écoval
Une boîte à outils pour accompagner les entreprises agro-alimentaires dans la réduction des consommations d’eau. AgroParisTech, Inrae, Prosim et cinq centres Actia (Actalia, Critt Paca, CTCPA, IFV, Iterg) viennent de terminer le projet ANR Minimeau (n° 17-CE10-0015) « Minimisation des consommations d’eau dans les industries agro-alimentaires par le développement d’une approche intégrée associant Empreinte eau et Pinch massique ».
Les résultats de ce projet démontrent un potentiel d’économies pouvant aller jusqu’à 30 %.
Une « boîte à outils Minimeau » avec sa vidéo explicative est mise à disposition des entreprises.
Contact : yvan.deloche@critt-iaa-paca.com
Avril 2022
![]() EQVEGAN : devenez un interlocuteur privilégié du projet sur les qualifications et compétences européennes pour l’industrie alimentaire végétalienne
EQVEGAN : devenez un interlocuteur privilégié du projet sur les qualifications et compétences européennes pour l’industrie alimentaire végétalienne
L’ACTIA et les partenaires du projet invitent les prestataires d’enseignement et de formation, les entreprises agro-alimentaires et autres acteurs, à devenir des interlocuteurs privilégiés du projet européen Erasmus+ Eqvegan. Ceux-ci pourront apporter leur expertise et bénéficier du développement de formations pour les étudiants et les professionnels de l’industrie agro-alimentaire végétalienne. Pour cela, rejoignez la base de données en remplissant le questionnaire. Un système de certification pour la reconnaissance des savoirs et des compétences des professionnels de l’alimentation végétalienne est en cours de développement.
Contact : Gemma Cornuau, g.cornuau@actia-asso.eu
Avril 2022
 BUDDIE-PACK : premier projet Horizon Europe gagné, pour l’ACTIA et ses centres, sur les emballages plastiques réutilisables
BUDDIE-PACK : premier projet Horizon Europe gagné, pour l’ACTIA et ses centres, sur les emballages plastiques réutilisables
Ce projet tirera parti de l’innovation comportementale, afin de promouvoir l’acceptation des emballages plastiques réutilisables (RPP) en analysant les moteurs et la psychologie de réutilisation des consommateurs. Il se concentrera sur des solutions de nettoyage innovantes, pour garantir la sécurité des consommateurs contre la contamination et le vieillissement éventuels des RPP, et sur la durabilité des propriétés fonctionnelles. Coordonné par IPC, centre partenaire de l’Actia, ce projet rassemblant vingt partenaires durera trois ans et demi, avec un budget d’environ 7 millions d’euros. L’Actia, ainsi que le LNE et le CTCPA, sont partenaires de Buddie-Pack.
Contact : Christophe Cotillon, c.cotillon@actia-asso.eu
Avril 2022
L’ACTIA et ses partenaires remportent deux projets européens Erasmus+ !
Le premier, e-SafeFoods, porte sur le développement d'un programme complet de formation en ligne sur la sécurité microbiologique dans les chaînes alimentaires, pour les professionnels actuels et futurs. Deux ITAI participent, Aerial et Actalia pour l’ACTIA, ainsi que trois partenaires européens : Alimento (Belgique), Institut polytechnique de Bragança (Portugal) et IsekiFood Association (Autriche). Le projet débutera en janvier 2022.
Le second, Fitness 2.0, prendra la suite du projet Fitness (septembre 2017 - août 2020), qui a permis de développer un logiciel pédagogique ouvert sur l’emballage alimentaire, pour l’enseignement supérieur et le personnel des entreprises. Ce projet permettra de continuer les travaux. Il implique l’ACTIA et le LNE, ainsi que neuf autres partenaires français et européens. Il devrait également débuter en janvier 2022.
Contact : Christophe Cotillon, c.cotillon@actia-asso.eu
Novembre 2021
 Retour sur la journée « prédiction des altérations microbiologiques et la biopréservation des aliments »
Retour sur la journée « prédiction des altérations microbiologiques et la biopréservation des aliments »
Le mardi 12 octobre 2021 a eu lieu une journée organisée par le RMT Actia Florepro et le consortium du projet ANR Redlosses pour restituer les travaux sur la prévision des altérations microbiologiques et la place de la biopréservation dans la conservation des aliments. Cette journée avait pour objectif d’informer l’ensemble des acteurs de l’agro-alimentaire des nouvelles connaissances générées dans le domaine de la microbiologie alimentaire, pour adapter les développements et les innovations aux attentes sociétales.
Retrouvez les vidéos et les présentations de la journée.
Contacts : souad.christieans@adiv.fr & monique.zagorec@inrae.fr
Novembre 2021
VIENT DE PARAÎTRE - Ouvrage « Agents antimicrobiens et sécurité sanitaire des aliments » aux éditions Lavoisier
Coordonné par le RMT Actia Chlean, l’ouvrage « Agents antimicrobiens et sécurité sanitaire des aliments » développe cette thématique en huit chapitres autour de la réglementation, des conservateurs et ingrédients à action antimicrobienne, des flores protectrices, auxiliaires technologiques, biocides désinfectants, de la validation de l’efficacité de la désinfection et des surfaces antimicrobiennes. Contact : Catherine Stride, cstride@ctcpa.org
Novembre 2021
 Projet européen Divinfood - Accroître la valeur des cultures négligées et sousutilisées dans les chaînes alimentaires – avec l’ACTIA et AERIAL
Projet européen Divinfood - Accroître la valeur des cultures négligées et sousutilisées dans les chaînes alimentaires – avec l’ACTIA et AERIAL
Coordonné par l’Inrae, ce projet européen comprenant vingt-six partenaires, dont l’Actia et Aerial, vient d’être accepté par la Commission, pour soutien après réévaluation. L’objectif global de ce projet multi-acteurs est de faciliter l’utilisation et accroître la valeur des cultures négligées et sous-utilisées (NUC) dans les chaînes alimentaires, pour favoriser des régimes alimentaires plus sains et des systèmes alimentaires plus durables. Pour y parvenir, Divinfood se concentrera sur des chaînes de valeur interactives de niveau court et intermédiaire qui peuvent répondre à la demande croissante des consommateurs, afin de produire des aliments plus sains à base de plantes ayant une identité locale/régionale, tout en promouvant des services et bénéfices durables pour l’agriculture et la transformation.
Divinfood étudiera des céréales et des légumineuses « mineures » dans trois régions géographiques confrontées à divers aléas climatiques et défis socio-économiques pour le développement de chaînes de valeur plus riches en agro-biodiversité.
Contact : Christophe Cotillon, c.cotillon@actia-asso.eu
Juillet 2021
 UMT Actia Agrobergo : les exosquelettes en agro-alimentaire
UMT Actia Agrobergo : les exosquelettes en agro-alimentaire
L’Adiv et Auvergne-Rhônes-Alpes Gourmand, partenaires de l’UMT Actia Agrobergo, multiplient les actions pour informer et accompagner les entreprises agro-alimentaires sur l’acquisition et l’intégration des exosquelettes. Ces actions sont réalisées dans le cadre du programme « Santé & performance » cofinancé par la Draaf/Dinaii et Auvergne-Rhônes-Alpes Gourmand, partenaire de la Carsat Rhône-Alpes depuis 2009 et de l’Adiv depuis 2017 dans le cadre de l’UMT Agrobergo. Récemment, les partenaires ont fait un inventaire des exosquelettes en agro-alimentaire. Téléchargez l’inventaire
Le 2 juillet 2021, un webinaire « Les exosquelettes dans l’agro-alimentaire. Pour réduire les efforts, l’exosquelette est-il une solution ? » a réuni plus de cent participants. Une thématique qui paraît prometteuse pour l’entreprise agro-alimentaire du futur !
Contacts : matthieu.alric@adiv.fr & Françoise Molegnana, f.molegnana@comite-arag.fr
Juillet 2021
![]()
L’ADIV et l’ACTIA partenaires du projet européen de coopération avec la Chine « DiTECT : Les TEChnologies Digitales comme facilitateur d'une transformation continue du système de sécurité sanitaire »
Projet réunissant 33 partenaires dont 21 de 10 pays européens et 12 de la Chine, Ditect a pour but de faciliter la création de chaînes d’approvisionnement alimentaire nationales et internationales transparentes et fiables, grâce à une approche intégrative soutenue par des technologies d’authentification et de traçabilité de pointe, utilisables sur l’ensemble de la chaîne alimentaire. Coordonné par l’université pour l’Agriculture d’Athènes, le projet a démarré le 1er novembre 2020 pour une durée de trente-six mois. L’Adiv est coresponsable avec un partenaire chinois d’un workpackage (WP2) visant à évaluer et suivre les risques de contamination tout au long de la chaîne alimentaire dans plusieurs secteurs et filières. L’ACTIA soutient l’Adiv dans ses actions de dissémination et communication au sein du projet (WP8).
Contact : Christophe Cotillon, c.cotillon@actia-asso.eu
Mai 2021

UMT Actia Protorisk : résultats du projet ANR Mobidic sur la contamination des ressources en eau par les parasites protozoaires
L’évaluation de la qualité biologique des ressources en eau repose sur des mesures d’indicateurs bactériens et nécessite le prélèvement de volumes d’eau importants. Les résultats de ces analyses dépendent, entre autres, du lieu et du moment où est prélevé l’échantillon, des paramètres physico-chimiques environnementaux ou encore des phénomènes météorologiques. Par ailleurs, les indicateurs bactériens ne reflètent pas la contamination en parasites protozoaires Cryptosporidium spp., Giardia duodenalis et Toxoplasma gondii, impliqués dans de nombreuses épidémies mondiales d’origine aquatique.
Le projet ANR Mobidic (2015-2020) mené dans le cadre de l'UMT proposait d’utiliser la capacité d’accumulation de deux espèces de mollusques bivalves, Dreissena polymorpha pour le domaine continental et Mytilus spp. pour le domaine côtier, pour évaluer la contamination des ressources en eau par les parasites protozoaires en question.
Ce projet a permis de démontrer la pertinence des bivalves comme outils alternatifs pour évaluer la contamination en parasites protozoaires des eaux continentales et côtières. Il ouvre de nouvelles perspectives dans le domaine de la surveillance de la qualité microbiologique des ressources en eau. Ce projet a obtenu le soutien du Feder.
Contact : Stéphanie La Carbona, s.lacarbona@actalia.eu
Avril 2021
![]() UMT Actia Newcarn : lancement du projet anémie-3D, une technologie innovante au service de la conception d’aliments mixtes
UMT Actia Newcarn : lancement du projet anémie-3D, une technologie innovante au service de la conception d’aliments mixtes
Ce projet porté par l'UMT vise à concevoir des aliments mixtes associant protéines animales et végétales par impression 3D. Destinés aux personnes souffrant d’anémie ferriprive, ces aliments mettent en jeu la complémentarité des sources de protéines :
- les produits carnés, notamment les abats, pour leur richesse en fer héminique biodisponible et leurs protéines de haute qualité nutritionnelle ;
- les légumineuses pour leur densité nutritionnelle et les propriétés texturantes de leurs protéines.
L’enjeu est de satisfaire les consommateurs d’un point de vue nutritionnel et organoleptique avec cette association singulière. Pour y parvenir, l’impression 3D alimentaire constitue une nouvelle voie d’innovation, qui offre une grande liberté de conception et permet ainsi l’élaboration d’aliments présentant une structure et une texture maîtrisées, ainsi qu’un aspect attrayant.
Contact : coline.schiell@adiv.fr et valerie.scislowski@adiv.fr
Avril 2021
Lancement de deux projets de formation européenS ERASMUS+
pour l'ACTIA
 PROJET EQVEGAN « Qualifications et compétences européennes pour l'industrie alimentaire végétalienne » le 1er décembre 2020
PROJET EQVEGAN « Qualifications et compétences européennes pour l'industrie alimentaire végétalienne » le 1er décembre 2020
Comprenant 15 partenaires de 11 pays, dont l’ACTIA, son objectif est de concevoir et proposer des formations innovantes sur l’alimentation végétalienne, à la fois aux professionnels et aux étudiants, en sept langues (anglais, croate, finnois, français, polonais, portugais et turc).
Un portail sur les compétences, qui comprendra une base de données des parties prenantes, des profils de professionnels ainsi qu’une base de données sur l’apprentissage sera développé. Un système européen de certification des profils de poste dans ce domaine sera élaboré, ainsi que des lignes directrices facilitant la reconnaissance des compétences, reposant sur des qualifications sectorielles identifiées, dans le but de favoriser la mobilité transnationale des professionnels dans ce secteur.
PROJET FOSTER « L’industrie alimentaire à l’ère dU numérique : nouvelles pratiques de formation et de développement des compétences, grâce à la réalité étendue » le 1er septembre 2020
L'objectif est d'introduire la réalité étendue (xR), qui permet de fusionner des éléments virtuels et réels dans un même environnement, dans le domaine de l'industrie alimentaire. Afin d'améliorer les conn![]() aissances et capacités des étudiants, chercheurs, universitaires et professionnels de l'industrie dans cette technologie innovante, les partenaires du projet développeront :
aissances et capacités des étudiants, chercheurs, universitaires et professionnels de l'industrie dans cette technologie innovante, les partenaires du projet développeront :
- deux programmes intensifs de formation ;
- un outil prototype xR adapté à l'industrie alimentaire ;
- un outil d'apprentissage en ligne sur l'utilisation de l'outil prototype xR, pour permettre le transfert de connaissances aux professionnels de l'industrie et aux universitaires.
Contact : Christophe Cotillon et Gemma Cornuau
Septembre 2020
PARUTION : INVENTAIRE Des méthodes et kits pour la détection des résidus de biocides désinfectants
 Dans ce document, les partenaires du RMT Actia Chlean ont recensé les différents tests et kits utilisables pour la détection de résidus de biocides, à partir des eaux de rinçage collectées après les opérations de nettoyage et désinfection. Il s'appuie notamment sur une étude menée dans le cadre du projet Resbiolact mené par le Cniel, l'Anses et Actalia.
Dans ce document, les partenaires du RMT Actia Chlean ont recensé les différents tests et kits utilisables pour la détection de résidus de biocides, à partir des eaux de rinçage collectées après les opérations de nettoyage et désinfection. Il s'appuie notamment sur une étude menée dans le cadre du projet Resbiolact mené par le Cniel, l'Anses et Actalia.
L'inventaire est téléchargeable gratuitement ici.
Septembre 2020
Il est maintenant possible de distinguer et tracer des souches de Bacillus thuringiensis du champ à l’assiette
Le 29 janvier 2020, avait lieu à Paris, la journée de restitution des projets nationaux BtID (financement Casdar) et Vegexpobt (financement FranceAgriMer) présentant différents outils permettant l’identification, la distinction et la traçabilité des souches commerciales de Bacillus thuringiensis utilisées au niveau européen pour leurs propriétés insecticides. Bacillus thuringiensis ou Bt est une bactérie ubiquiste utilisée dans le domaine agricole en protection des cultures dans les années 1950. C’est l’insecticide le plus utilisé au monde en agriculture biologique. Au niveau taxonomique, Bt appartient au groupe Bacillus cereus et reste difficile à différencier au sein de ce groupe, ce qui peut entraîner des toxi-infections alimentaires collectives ou des altérations microbiologiques.
À l’heure actuelle, B. cereus ne fait pas l’objet de critères sanitaires selon la réglementation européenne. Cependant, l’utilisation de ces bio insecticides engendre des niveaux de contamination, qui peuvent impacter fortement les industries agro-alimentaires fabriquant des aliments à base de végétaux traités par Bt. La table ronde a permis d’appréhender la problématique de manière transversale en soulignant le besoin de légiférer et statuer sur :
- l’innocuité des souches commerciales de Bt autorisées au niveau européen, qui ont pu être associées mais non impliquées, à des cas d’intoxication ;![]()
- le manque d’alternative aux pesticides chimiques si l’utilisation de bio insecticides à base de Bt était compromise, bien que ce soit une solution biologique efficace, avantageuse et durable pour la protection des cultures.
Coordinateurs : Florence Postollec, UMT Actia Alteri’x, florence.postollec@adria.tm.fr rodolphe.vidal@itab.asso.fr - Catherine Denis, c.denis@actalia.eu
Juillet 2020
Accroître les compétences dans le domaine de l’efficacité énergétique pour une industrie agro-alimentaire plus durable grâce au projet Induce![]()
Le projet européen Induce a permis le développement d’une plate-forme facilement accessible (boîte à outils Induce) où des formations, des cours en ligne, des lignes directrices et outils sont mis à la disposition des entreprises désirant améliorer leur efficacité énergétique. Cette plate-forme intègre également la méthodologie Induce : un programme spécifique d’acquisition de compétences, pour chaque entreprise accédant à la boîte à outils, a été développé. Cela
permet ainsi de trouver le programme ad hoc adapté aux besoins et possibilités de l’entreprise.
La méthodologie Induce fait le lien entre transfert de connaissances sur l’efficacité énergétique et changement des modèles organisationnels et comportementaux, permettant ainsi un vrai changement de culture et d’environnement, au sein même de l’entreprise et de ses salariés.
Site internet Induce- compte Twitter
Contact : Christophe Cotillon et Gemma Cornuau
Juillet 2020
Le projet Enabling met à disposition des outils pour soutenir les réseaux locaux d’innovation sur la biomasse![]()
Depuis 2017, les partenaires du projet Horizon 2020 Enabling œuvrent pour créer des conditions appropriées au développement de biomasse utile pour les chaînes de valeur des BBPs (produits et procédés d’origine biologique). Enabling a ainsi développé plusieurs plate-formes facilitant la communication et améliorant le développement et la transformation de la biomasse :
- un atlas des bonnes pratiques permet de collecter les meilleures pratiques des BBPs, en Europe et au-delà, qui sont entièrement ou partiellement transférables à d’autres régions, ou bien qui servent d’inspiration pour les partenaires de la chaîne de valeur ;
- des activités d’accompagnement sont disponibles via un simple questionnaire en ligne, afin d’aider les producteurs de biomasse ou l’industrie BBP à adopter les meilleures pratiques émergentes et de permettre aux candidats de mieux cerner les caractéristiques de la bioéconomie et de ses composantes.
Contact : Christophe Cotillon et Gemma Cornuau
Juillet 2020
FIELDS : NOUVeau projet européen de formation pour l'actia![]()
Le projet Fields, financé par le programme Erasmus+, a été lancé les 3 et 4 février 2020 à Turin (Italie). Coordonné par l’université de Turin, il comprend trente partenaires dont l’ACTIA, de douze pays différents et durera quatre ans. Il vise à répondre aux besoins actuels et futurs de compétences de l’agriculture européenne sur les aspects de durabilité, de numérique et de bioéconomie.
Fields conduira ainsi à l’élaboration d’une stratégie européenne, qui sera définie sur la base d’une analyse approfondie des besoins et sera mise en pratique dans le projet, à travers la création de guides et méthodes, ainsi que de programmes et de contenus de formation.
Contact : Christophe Cotillon et Gemma Cornuau
Avril 2020
Lancement du projet de recherche Casdar « DiVinCidre » :
![]()
Développement d'Itinéraires techniques pour optimiser le caractère fruité des vins et des cidres (2020-2022)
À ce jour, peu de paramètres de qualité des moûts permettent aux professionnels de prévoir de façon certaine le profil aromatique des vins et des cidres, même lorsque les itinéraires techniques sont parfaitement maîtrisés.
L’objectif du projet DiVincidre est de mettre à disposition des élaborateurs de cidres et de vins, les éléments qui leur permettent de choisir les itinéraires de production adéquats pour mieux maîtriser le profil organoleptique des produits finis.
L’approche proposée dans ce projet est de prendre en considération plusieurs facteurs nutritionnels des levures afin de mieux expliquer la formation des composés aromatiques marqueurs du « fruité ». La mise en évidence des facteurs nutritionnels les plus impactants sur le « fruité » des cidres et des vins, doit permettre de proposer un diagnostic analytique pertinent du « potentiel aromatique fruité » des moûts, mais aussi de sélectionner des itinéraires techniques d’élaboration les favorisant.
Partenaire du projet : IFPC (pilote), IFV, Inrae Montpellier, unité Science pour l’œnologie (SPO), Nyseos, en collaboration avec l’UMT Actia Nova²Cidre et le RMT Actia Quadiferm
Contact : pascal.poupard@ifpc.eu
Février 2020
 Déclaration nutritionnelle obligatoire : une méthode rapide pour faciliter les analyses de composition dans la viande de porc
Déclaration nutritionnelle obligatoire : une méthode rapide pour faciliter les analyses de composition dans la viande de porc
Pour faciliter l’étiquetage nutritionnel dans la filière porc, l’Ifip travaille sur des méthodes rapides de mesure des matières grasses, acides gras saturés, protéines et humidité. La spectroscopie proche infrarouge (NIRS) semble une bonne alternative aux méthodes d’analyses chimiques.
Pour évaluer efficacement les performances de la méthode, l’Ifip a tout d’abord construit des bases de données spécifiques présentant une surreprésentation des valeurs extrêmes pour le rapport lipides/protéines et dont la composition des lipides a été modifiée par la mise en place de plusieurs modalités d’alimentation. Les équations de prévision mises au point ont été ensuite validées sur des données issues de la filière porcine pour en évaluer la précision.
La validation externe des calibrations NIRS développées a ainsi de très bonnes performances pour la prévision des taux de lipides, d’humidité et de protéines de la viande. De plus, les modèles de prévision des quantités d’acides gras saturés montrent de bonnes performances lorsqu’exprimées en quantités pour 100 g de tissu, conformément aux exigences du règlement (UE) n° 1169/2011 dit « Inco ».
Étude financée par FranceAgrimer.
Contact : antoine.vautier@ifip.asso.fr
Février 2020

Le projet Ecoqualina « Écoconception des produits sous signe de qualité de Nouvelle-Aquitaine », une action fédératrice pour le Réseau Actia Nouvelle-Aquitaine
 Le réseau ACTIA Nouvelle-Aquitaine et l’Agence de l’Alimentation Nouvelle-Aquitaine (Aana) ont initié un projet pour accompagner des filières en Nouvelle-Aquitaine sous Signe officiel de la qualité et de l’origine (SIQO) dans une démarche d’éco-conception.
Le réseau ACTIA Nouvelle-Aquitaine et l’Agence de l’Alimentation Nouvelle-Aquitaine (Aana) ont initié un projet pour accompagner des filières en Nouvelle-Aquitaine sous Signe officiel de la qualité et de l’origine (SIQO) dans une démarche d’éco-conception.
Il vise à identifier, en vue de leur éventuelle intégration aux côtés des critères de qualité spécifiques au produit, des critères d’amélioration de la performance environnementale du produit tout au long de sa chaîne de valeur.
Quinze produits seront étudiés : agneau de Poitou-Charentes IGP/LR, agneau du Limousin IGP - agneau du Périgord IGP/LR, agneau de lait des Pyrénées IGP/LR, fromage Ossau-Iraty AOP, beurre Charentes-Poitou AOP, jambon de Bayonne IGP, porc du Sud-Ouest IGP, pruneau d’Agen IGP, huîtres Marennes Oléron IGP/LR.
* IGP : Indication géographique protégée - LR : Label rouge - AOP : Appellation d’origine protégée
Contact : Fabrice Bosque, f.bosque@iterg.com
Février 2020
LES PARTENAIRES DU PROJET PRO-ORG RECHERCHENT DES ENTREPRISES POUR TESTER LE GUIDE SUR LA TRANSFORMATION DES PRODUITS BIO 
L’université des Sciences de la vie de Varsovie (Pologne) partenaire du projet Core Organic Pro-Org visant à proposer un guide de bonnes pratiques de transformation à l’usage des opérateurs bio européens, a organisé une réunion de projet, les 11 et 12 septembre 2019, à Varsovie. À cette occasion, les partenaires ont pu travailler sur la proposition de cadre d’évaluation des procédés de transformation, présenter les résultats de l’étude « Bio produit » menée par l’université de Münster et planifier le lancement de l’étude marketing par le FIBL (Institut de recherche de l’agriculture biologique de Suisse).
Le projet arrive à mi-parcours, les partenaires sont actuellement à la recherche d’entreprises qui souhaiteraient participer à la phase d’essai du guide de bonnes pratiques.
Concrètement, il s’agira de tester le cadre d’évaluation afin de vérifier sa pertinence pour différentes filières et s’il correspond aux attentes des entreprises afin d’améliorer les différentes qualités de leurs produits transformés.
Pour participer au test, merci de contacter Rodolphe Vidal : rodolphe.vidal@itab.asso.fr ou Christophe Cotillon, c.cotillon@actia-asso.eu.
Pour suivre le projet, vous pouvez vous inscrire à la lettre d’information Pro-Org en bas des pages du site.
Décembre 2019

DES FLORES PROTECTRICES POUR DÉFENDRE LES SURFACES DES ATELIERS CHARCUTIERS CONTRE LES GERMES INDÉSIRABLES
Dans ce projet Prosurf mené par l’Ifip, AgroParisTech et la société Chr.Hansen et financé par FranceAgrimer, l’objectif est de mettre au point un procédé d’aspersion de flores protectrices sur des surfaces ouvertes, puis d’évaluer les performances de deux ferments à l’encontre de l’implantation de flores indésirables. Deux germes ont été choisis, un germe pathogène (Listeria monocytogenes) et un germe d’altération (Pseudomonas fluorescens), ainsi que deux types de matériaux de surface (acier inoxydable et polymère).
Les différents essais ont permis de valider les paramètres : type de buse, temps de pulvérisation, cycle de travail, distance de pulvérisation, temps de séchage. Les essais de co-inoculation des ferments et des germes indésirables ont montré l’efficacité d’un ferment spécifique sur Listeria monocytogenes.
Des essais complémentaires sont en cours pour renforcer l’adhérence des ferments aux matériaux et ainsi optimiser l’efficacité barrière à l’encontre de l’implantation des flores indésirables. Des essais sur ligne pilote seront ensuite programmés.
Contact : bastien.fremaux@ifip.asso.fr
Octobre 2019
Projet Fitness : la plate-forme digitale sur les emballages en test !![]()
Le projet Fitness, coordonné par l’Actia assistée du LNE et financé dans le cadre du programme européen Erasmus+, a pour but de mettre en place, d’ici août 2020, une plate-forme digitale ouverte à tous sur la thématique de l’emballage. Elle contiendra, entre autres, des cours, des guides, des quiz et des webinaires en anglais adaptés à différents niveaux de compétences.
Depuis l’été 2019, cette plate-forme en ligne existe sous forme beta et une partie des cours est déjà consultable. La prochaine phase du projet consiste à tester les contenus auprès de publics cibles, étudiants, doctorants, enseignants et industriels dans les différents pays partenaires du projet : Allemagne, Croatie, Espagne, France, Portugal. Les commentaires et retours collectés à l’issue de ces sessions permettront d’améliorer les contenus.
La première session organisée à Freising (Allemagne) par TUM / Fraunhofer pendant trois jours, du 9 au 11 septembre 2019, a été un véritable succès !
Contacts : Gemma Cornuau, g.cornuau@actia-asso.eu & catherine.loriot@lne.fr
Octobre 2019
 IPC MONTE EN CAPACITÉ SUR LE RECYCLAGE DES EMBALLAGES EN PLASTIQUE : DEUX PROJETS DE RECHERCHE AVEC CITEO ET D'AUTRES PARTENAIRES
IPC MONTE EN CAPACITÉ SUR LE RECYCLAGE DES EMBALLAGES EN PLASTIQUE : DEUX PROJETS DE RECHERCHE AVEC CITEO ET D'AUTRES PARTENAIRES
SUPErPE, porté par un industriel du film et en partenariat avec le CTCPA, a pour objectif de développer une technologie de recyclage du polyéthylène et du polypropylène basée sur l’utilisation du CO2 supercritique. Il associe également un industriel de l’emballage cosmétique, une unité de recherche de l’université de Saint-Étienne et une plate-forme spécialisée dans la technologie supercritique.
Le deuxième projet, conclu avec le Cotrep (Centre de ressources et d’expertise sur la recyclabilité des emballages ménagers en plastiques), réunit Citeo, Valorplaste et Elipso et a pour objectif d’améliorer la recyclabilité des emballages souples. Une ligne de recyclage test de petite capacité sera mise en place sur deux des sites d’IPC, à Oyonnax et Alençon. Sur la base d’un protocole établi avec les deux recycleurs français d’emballages souples, Barbier et Machaon, elle reproduira les étapes du recyclage (broyage, lavage etc.) pour tester et étudier le comportement de ces emballages chez les recycleurs.
Ces deux projets portent sur deux catégories d’emballages souples, les films plastiques étirables en polyéthylène (PE) et les complexes constitués de plusieurs composants et / ou d’autres types de résines plastiques, comme le polypropylène (PP).
Contact : jacques.thebault@ct-ipc.com
Octobre 2019
Projet Foodfreeze : comment gérer le givre dans les entreprises agro-alimentaires ?
 La gestion du givre, lors des opérations de surgélation et de stockage des produits, est faite de manière empirique. Or le givre a un impact sur l’efficacité énergétique des installations, mais potentiellement aussi sur la qualité des produits congelés ou surgelés.
La gestion du givre, lors des opérations de surgélation et de stockage des produits, est faite de manière empirique. Or le givre a un impact sur l’efficacité énergétique des installations, mais potentiellement aussi sur la qualité des produits congelés ou surgelés.
Quatre partenaires du Réseau Actia Écofluides, Oniris (coordinateur), le Cetiat, Aquimer et le Critt Agroalimentaire Paca se sont donc saisis de ce sujet au côté d’un fournisseur d’équipements, GEA Réfrigération. Retenu à l’appel à projets de recherche Énergie durable de l’Ademe, Foodfreeze a démarré en juin 2018, et se terminera fin 2021. Foodfreeze permettra de proposer aux industriels de l’agro-alimentaire et aux professionnels du froid des modèles numériques de simulation de la croissance du givre afin d’optimiser les séquences de dégivrage. Une méthode de diagnostic des surgélateurs sera également développée.
Contact : cyril.toublanc@oniris-nantes.fr
Juillet 2019
Développement des circuits courts : votre avis nous intéresse pour faire avancer le projet européen Smartchain
 L’ACTIA et le CTCPA participent au projet européen Smartchain sur les circuits courts qui rassemble quarante-trois partenaires issus de onze pays. Il se base sur dix-huit études de cas de circuits courts réparties dans toute l’Europe. L’objectif principal du projet est d’identifier, à partir de ces cas d’études, les facteurs clés permettant d’influencer le développement du circuit court.
L’ACTIA et le CTCPA participent au projet européen Smartchain sur les circuits courts qui rassemble quarante-trois partenaires issus de onze pays. Il se base sur dix-huit études de cas de circuits courts réparties dans toute l’Europe. L’objectif principal du projet est d’identifier, à partir de ces cas d’études, les facteurs clés permettant d’influencer le développement du circuit court.
Dans le cadre de ce projet, un questionnaire a été élaboré afin d’identifier les leviers et obstacles au développement de ces circuits. Il est destiné à tous les acteurs de la chaîne agro-alimentaire que ce soient les producteurs, transformateurs, distributeurs, mais aussi toutes les structures de support et d’accompagnement.
Merci de prendre quelques minutes pour répondre au questionnaire disponible via ce lien. Vos connaissances et votre retour d’expérience nous apporteront une aide précieuse pour faire avancer le projet.
Ce projet est financé par le programme de recherche et d’innovation Horizon 2020 de l’Union européenne dans le cadre de la convention de subvention n° 773785.
Juillet 2019
Le projet Oxyvir de l’UMT Actia Virocontrol mis à l’honneur au salon de l’Agriculture
 Vendredi 1er mars 2019 au salon de l’Agriculture s’est tenue une table ronde sur le thème « Les aquacultures, des filières d’avenir », organisée par la direction des Pêches maritimes et de l’Aquaculture du ministère de l’Agriculture. Actalia y a été convié pour présenter le projet Oxyvir, financé par le FEAMP et la région Normandie, qui a pour but de développer une méthode permettant de discriminer les norovirus infectieux des norovirus non infectieux dans les coquillages, et ainsi améliorer la gestion du danger à norovirus par les pouvoirs publics. Choisi parmi une cinquantaine de projets financés par le FEAMP, les résultats d’Oxyvir attendus pour 2020 représentent un réel enjeu pour l’aquaculture en participant à la pérennité de la filière conchylicole.
Vendredi 1er mars 2019 au salon de l’Agriculture s’est tenue une table ronde sur le thème « Les aquacultures, des filières d’avenir », organisée par la direction des Pêches maritimes et de l’Aquaculture du ministère de l’Agriculture. Actalia y a été convié pour présenter le projet Oxyvir, financé par le FEAMP et la région Normandie, qui a pour but de développer une méthode permettant de discriminer les norovirus infectieux des norovirus non infectieux dans les coquillages, et ainsi améliorer la gestion du danger à norovirus par les pouvoirs publics. Choisi parmi une cinquantaine de projets financés par le FEAMP, les résultats d’Oxyvir attendus pour 2020 représentent un réel enjeu pour l’aquaculture en participant à la pérennité de la filière conchylicole.
Contact : Nicolas Boudaud, n.boudaud@actalia.eu
Mars 2019
L’Actia édite chaque trimestre une lettre d’information scientifique et technique faisant une synthèse des activités et résultats scientifiques des centres du réseau Actia. Vous pouvez télécharger ci-dessous toutes les lettres Actialités ayant été publiées.
ABONNEMENT LETTRE D'INFORMATION ACTIA

POUR ACCÉDER AUX REVUES DE PRESSE VEUILLEZ VOUS IDENTIFIER
RÉSERVÉ AUX MEMBRES DU RÉSEAU ACTIA AYANT DÉJÀ REÇU
LEURS IDENTIFIANTS D’ACCÈS EXTRANET.




